Effets biologiques des rayonnements ionisants
Interaction des rayonnements avec la matière vivante
Effets cellulaires
Courbes de survie et leur signification
Courbe exponentielle (fig. 5-1)
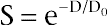
On constate que le nombre de cellules survivantes diminue de façon exponentielle avec la dose. En coordonnées semi-logarithmiques, cette courbe est représentée par une droite de pente α , qui exprime la radiosensibilité de la population cellulaire. On utilise pour exprimer cette sensibilité la dose D0, égale à l’inverse de la pente, 1/ α , qui est donc la dose qui laisse survivre 37 % des cellules. La dose qui tue la moitié des cellules est appelée « dose létale 50 » (DL50). Elle est égale à D0 × Log2 (soit 0,693D0).
Courbe sigmoïde (fig. 5-2)
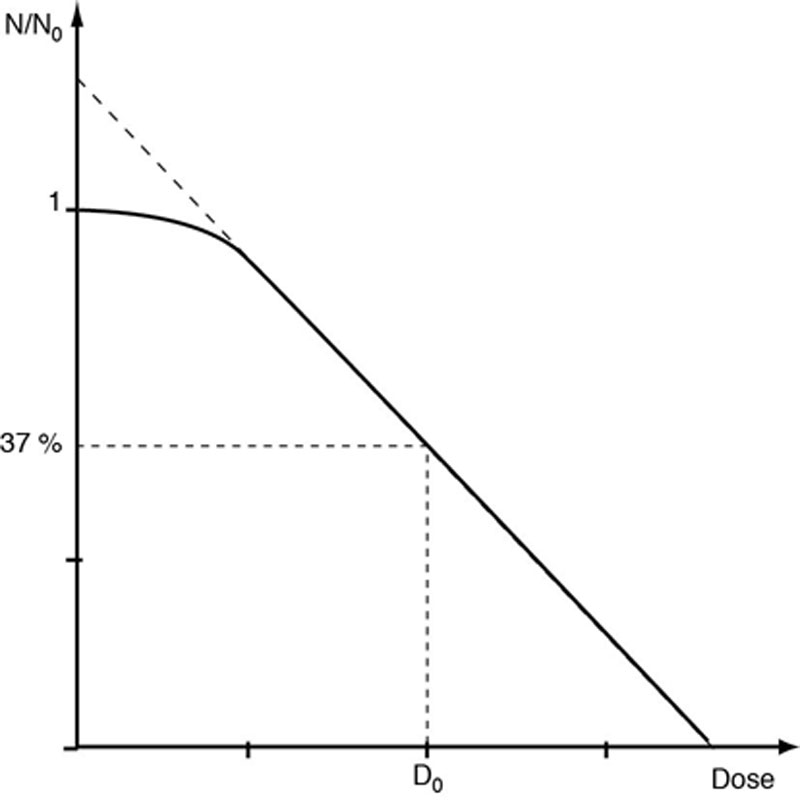
Elle s’observe avec la plupart des cellules de mammifères. La partie initiale de la courbe n’est plus rectiligne en coordonnées semi-logarithmiques. D0 n’est calculable qu’ à partir de cette partie rectiligne, elle augmente donc par rapport à la courbe exponentielle. Cette courbe traduit la capacité de réparation des cellules aux faibles doses.
Effets déterministes
Effets aléatoires ou stochastiques
Effets moléculaires
Lésions de l’ADN
Related posts:
 Grossesse et exposition médicale
Grossesse et exposition médicale
 Effets pathologiques des rayonnements ionisants : effets stochastiques
Effets pathologiques des rayonnements ionisants : effets stochastiques
 Organisation de la radioprotection
Organisation de la radioprotection
 Interaction des rayonnements ionisants avec la matière
Interaction des rayonnements ionisants avec la matière
 Doses délivrées au patient en radiodiagnostic. Quantification et optimisation
Doses délivrées au patient en radiodiagnostic. Quantification et optimisation
![]() Exposition naturelle et artificielle
Exposition naturelle et artificielle
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Effets biologiques des rayonnements ionisants
5
Les premiers effets cliniques attribuables aux rayonnements ionisants (RI) ont été rapportés très tôt après la découverte des rayonnements X et de la radioactivité à la fin du XIXe siècle. La première radiolésion a été décrite en 1896, les premiers effets tératogènes en 1901 et le premier cancer humain radio-induit en 1902. La cancérogenèse était démontrée expérimentalement en 1910. La multiplicité de ces conséquences cliniques a ouvert la voie à la radiobiologie, étude des mécanismes aboutissant à ces lésions, afin de les prévenir.
L’unité élémentaire d’un tissu vivant est la cellule. Ce chapitre sera consacré à la description des effets des rayonnements à l’ échelle cellulaire dans un premier temps, à l’ échelle moléculaire dans un deuxième temps. Ce second niveau d’ étude permettra de comprendre les effets cellulaires.
Les rayonnements ionisants interagissent avec les atomes constituant la matière vivante comme avec les atomes de la matière inorganique. Les mécanismes d’interaction ont été présentés en détail au chapitre 2. Une notion fondamentale à rappeler pour bien percevoir l’effet des rayonnements dans les milieux biologiques est le faible transfert énergétique concerné. Ainsi, une dose de 1 gray absorbée dans un tissu correspond par définition à une énergie de 1 Joule transférée à un kilogramme de matière. En termes d’ énergie dissipée sous forme thermique dans un milieu aqueux, cela équivaut à une élévation de température d’un quart de millième de degré Celsius. Ceci signifie que l’effet thermique, très faible, ne contribue en rien aux conséquences d’une telle dose, pourtant responsable d’effets biologiques significatifs. Les effets biologiques sont donc d’origine biochimique, conséquence des ionisations produites au sein du milieu irradié, sans participation d’une composante physique.
De même, cette dose de 1 gray produit une ionisation pour 100 millions de molécules d’eau. Ceci correspond à une augmentation du nombre d’ions apparaissant spontanément dans une cellule d’un ion supplémentaire pour 100 000 ions « naturels ». L’effet des rayonnements est donc, là encore, quantitativement minime. Cependant, les ions produits conduisent à la formation d’espèces chimiques très réactives. Il s’agit de radicaux libres, structures chimiques comportant un électron périphérique non apparié à l’origine de ces propriétés. Ainsi, le radical hydrogène, noté H•, est-il puissamment réducteur et le radical hydroxyle, HO•, très fortement oxydant. La formation de ces espèces sera précisée plus loin dans la description du processus de radiolyse de l’eau.
Les ionisations engendrées par un rayonnement sont primaires, produites par le rayonnement lui-même, ou secondaires, induites par les électrons mis en mouvement. Elles sont distribuées le long de la trajectoire du rayonnement. Le paramètre majeur est, nous l’avons vu, la densité des interactions le long de la trajectoire du rayonnement dans le milieu. Elle permet de définir le transfert linéique d’ énergie (TLE), caractéristique d’un rayonnement donné pour une énergie donnée. Ainsi, les interactions d’un rayonnement présentant un TLE élevé avec une structure moléculaire ont un risque élevé de lésions de celle-ci. À dose absorbée égale, les effets biologiques d’un rayonnement à TLE élevé seront donc potentiellement beaucoup plus importants que ceux d’un rayonnement à faible TLE.
Cette propriété est à l’origine de la notion de facteur de qualité des rayonnements qui tient compte de la différence de TLE existant entre les rayonnements électromagnétiques et les différentes particules (cf. chapitre 3). Pour un effet donné, on peut aussi faire le rapport entre l’ énergie nécessaire d’un rayonnement donné pour obtenir cet effet et l’ énergie nécessaire avec un autre rayonnement pour obtenir le même effet. Ce rapport est connu sous le nom d’effet biologique relatif des rayonnements (EBR). Il est défini spécifiquement pour chaque effet.
Les effets élémentaires sur la cellule ont été pressentis dès l’identification des effets biologiques macroscopiques des rayonnements ionisants. En 1906, Bergonié et Tribondeau ont établi une relation entre la radiosensibilité des cellules et leur activité mitotique, leur devenir caryocinétique (nombre de mitoses programmées dans la vie du sujet) et leur différenciation. Plus une cellule se divise souvent, plus le nombre de divisions qu’elle doit accomplir est grand, moins elle est différenciée et plus elle est radiosensible. En corollaire, plus une cellule est différenciée, moins souvent et moins longtemps elle se multiplie et plus elle est radiorésistante. C’est cette loi qui explique qu’au sein d’un tissu irradié, ce sont les cellules souches non différenciées, dont la tâche est de fournir par multiplication les cellules qui vont secondairement se différencier pour construire le tissu, qui sont les premières éliminées. Cette loi est également à la base de toutes les applications des RI dans le traitement des affections néoplasiques puisque la capacité de prolifération des cellules malignes est supérieure à celle des cellules normales et les affecte donc d’une radiosensibilité plus élevée.
Les expériences d’irradiation de cultures cellulaires ont permis de construire les premières théories relatives aux mécanismes d’action des RI sur les cellules et de fournir des évaluations quantitatives des effets observés. En fonction de la forme des courbes de survie, il est possible de déduire plusieurs types de comportement cellulaire.
Le modèle exponentiel (ou linéaire en représentation semi-logarithmique), chez les bactéries, répond à une équation simple :
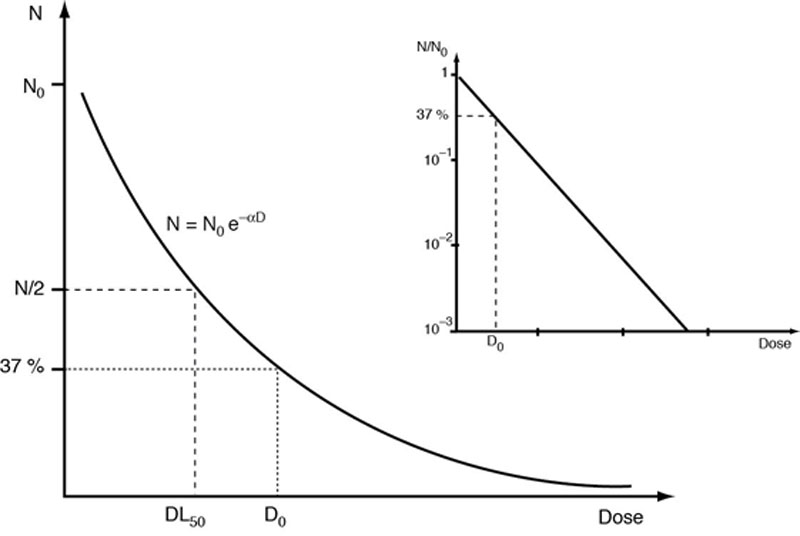
où S est la survie c’est-à-dire le rapport du nombre de cellules survivantes sur le nombre initial de cellules, N/N0 et D0 la dose pour laquelle, il reste 37 % de cellules survivantes, il s’agit alors de modèles à une cible létale et un impact est suffisant pour détruire une cellule, selon un modèle aléatoire.
Figure 5-1 Courbe de survie cellulaire exponentielle.
Ce modèle avec épaulement, en représentation semi-logarithmique, correspond à un cas de figure plus complexe, rencontré pour les cellules de mammifères. Il s’explique non plus par le modèle à plusieurs cibles sublétales, non étayé par les données radiobiologiques modernes mais par la mise en évidence des systèmes de réparation cellulaire à faible dose.
Figure 5-2 Courbe de survie avec épaulement.
Ces courbes modélisent la mort cellulaire induite par les RI, qui est à l’origine des effets déterministes des rayonnements. Ces effets apparaissent au-dessus d’un seuil, dépendant de l’effet et du tissu considéré, et leur gravité augmente ensuite avec la dose. Le seuil d’apparition des symptômes cliniques est relié au pourcentage de cellules d’un tissu dont la mort est nécessaire pour désorganiser suffisamment sa fonction. La gravité clinique augmente avec la dose délivrée au tissu puisque le pourcentage de cellules tuées augmente lui aussi avec cette dose. Enfin, pour un tissu donné, il existe toujours un intervalle libre entre l’irradiation et l’apparition des symptômes ; ce délai est lié d’une part à la durée de vie des cellules différenciées, moins radiosensibles que les cellules souches et d’autre part à la mort cellulaire différée avant laquelle se produisent quelques divisions des cellules souches ; lorsque la dose augmente, la fraction de mort immédiate augmente et l’accélération de l’ élimination des cellules souches est corrélée à la gravité de l’ état clinique observé à la phase d’ état.
Les RI peuvent aussi léser la cellule sans la tuer, en créant des altérations cellulaires compatibles avec la survie cellulaire mais potentiellement néfastes à plus ou moins long terme. Les effets de ce type les plus importants sont les modifications chromosomiques. Ces effets relèvent d’un phénomène statistique au sein de la population cellulaire car ils surviennent au hasard dans les cellules. Ce sont des effets non déterministes, ou stochastiques des RI. Ils sont de deux types en fonction de la nature de la cellule concernée : effet cancérogène lorsque la cellule concernée est une cellule somatique et effet héréditaire transmissible lorsque la cellule est de nature germinale. Les effets stochastiques s’opposent en tous points dans leurs caractéristiques aux effets déterministes.
• Ils ne sont pas spécifiques : un cancer ou une mutation transmissible induit par les RI est indiscernable d’une affection spontanée naturelle.
• Leur gravité est sans lien avec la dose car une fois déclenchées, ces affections évoluent pour leur propre compte avec le même génie évolutif que les affections naturelles.
• Leur probabilité de survenue croît avec la dose jusqu’ à un maximum où le nombre de cellules tuées devient trop important pour laisser suffisamment de cellules susceptibles d’ être à l’origine d’une affection cancéreuse.
• Ils n’ont, en théorie, pas de seuil de survenue car un seul impact dans une cellule pourrait être à l’origine d’un cancer ; en réalité la situation est beaucoup plus complexe et il n’existe pas à ce jour de démonstration, épidémiologique ou biologique, de ce postulat qui est à la base de l’utilisation de la relation linéaire sans seuil pour calculer les effets des faibles doses de RI.
On voit donc que les effets des RI à l’ échelle cellulaire ne sont pas univoques. Il est nécessaire d’envisager l’ échelle moléculaire pour trouver des éléments de réponse à la diversité des effets observés.
Au sein d’une cellule peuvent être identifiées plusieurs cibles potentielles : la membrane cellulaire, les organites cytoplasmiques et le noyau. Les deux premières de ces structures ne sont concernées que par les doses les plus fortes qui conduisent dans tous les cas à la mort cellulaire. C’est l’atteinte du noyau, plus particulièrement de son élément le plus noble, la molécule d’ADN porteuse du code génétique de la cellule, qui rend compte des effets stochastiques.
La molécule d’ADN présente de multiples particularités et l’une des plus intéressantes est l’existence d’une machinerie enzymatique très complexe dédiée au maintien de son intégrité et à la réparation la plus fidèle possible d’ éventuelles lésions. Cet équipement enzymatique est fondamental pour la survie cellulaire, ce qui explique qu’il soit très constant pour l’ensemble des cellules et organismes vivants depuis les bactéries jusqu’aux mammifères. Ceci a permis d’extrapoler de nombreuses observations effectuées sur des organismes simples aux cellules de mammifères. Les lésions de l’ADN peuvent procéder de nombreuses causes, intrinsèques à la cellule ou extrinsèques. La plus importante est certainement liée à la production de radicaux libres au cours du métabolisme cellulaire de l’oxygène. Ce dernier peut donc être considéré comme indispensable à la vie cellulaire mais également comme la source des espèces chimiques les plus toxiques pour l’ADN. Les RI font partie des agresseurs extrinsèques potentiels, au même titre que de multiples toxiques chimiques, souvent plus efficaces pour induire des anomalies de l’ADN.
Les RI les provoquent par l’intermédiaire des ionisations moléculaires. Il est important de préciser d’emblée que les lésions provoquées par les rayonnements ne sont pas spécifiques. Ainsi, les lésions qui vont être décrites peuvent survenir spontanément dans une cellule ou être induites par d’autres toxiques. Il n’est donc pas possible d’affirmer la responsabilité de RI dans la genèse de lésions présentes dans une cellule. Il n’existe qu’un élément d’orientation lié au mode d’action des rayonnements, à savoir le caractère localement concentré de lésions sur un segment de l’ADN. Cette densité de lésions peut être expliquée par la localisation des interactions causales sur la trajectoire du rayonnement surtout en cas de TLE élevé.
