14 Systèmes Sensoriels
14.1 Afférences somatosensories à la moelle spinale
14.2 Réflexes et voies médullaires
14.3 Système somatosensoriel : les voies spinocérébelleuses
14.4 Système somatosensoriel : les colonnes dorsales et les modalités épicritiques
14.5 Système somatosensoriel : les voies spinothalamiques et spinoréticulaires et les modalités protopathiques
14.6 Traitement de la douleur par les voies spinothalamiques et spinoréticulaires de la moelle spinale
14.7 Mécanismes de la douleur neuropathique et de la douleur continue sympathique
14.8 Contrôle descendant des systèmes somatosensoriels ascendants
14.9 Systèmes sensoriels trigéminés et associés
14.10 Anatomie des papilles gustatives et de leurs récepteurs
14.12 Voies périphériques de la perception du son
14.13 Labyrinthes osseux et membraneux
14.14 Innervation des cellules ciliées de l’organe de Corti par le VIII
14.16 Voies auditives afférentes
14.17 Voies auditives centrifuges (efférentes)
14.18 Récepteurs vestibulaires
14.22 Chambres antérieure et postérieure de l’œil
14.23 Rétine : les couches rétiniennes
14.24 Rétine : les photorécepteurs
14.25 Rétine : le nerf optique
14.26 Artères et veines de l’œil
14.27 Anatomie et relations avec le chiasma optique
14.28 Voies visuelles : projections rétiniennes vers le thalamus, l’hypothalamus et le tronc cérébral
14.29 Voies visuelles : la voie réticulo-géniculo-calcarine
14.30 Voies visuelles dans les lobes pariétal et temporal
Systèmes somatosensoriels
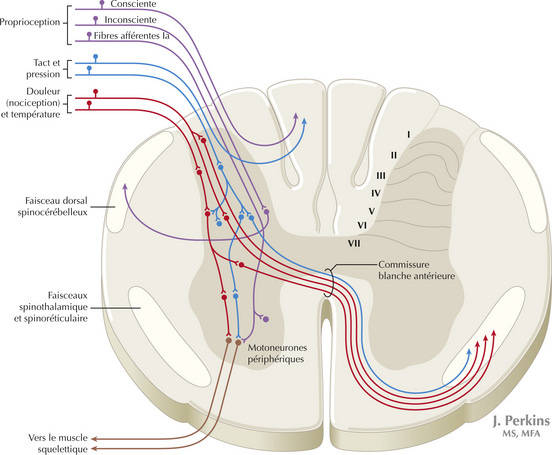
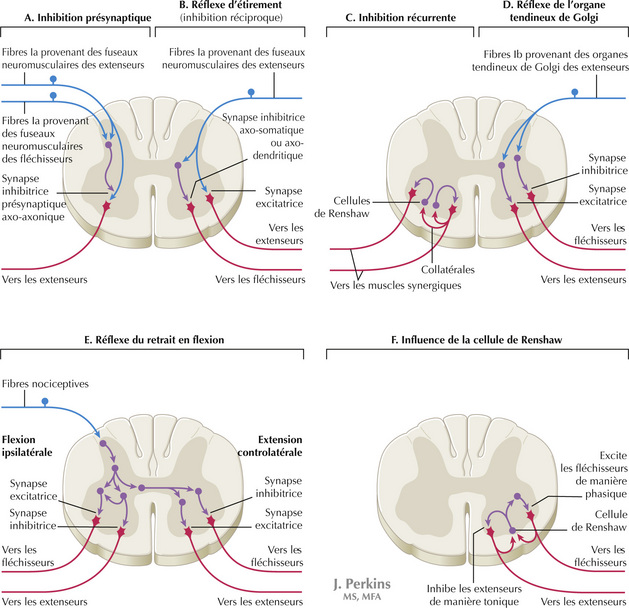
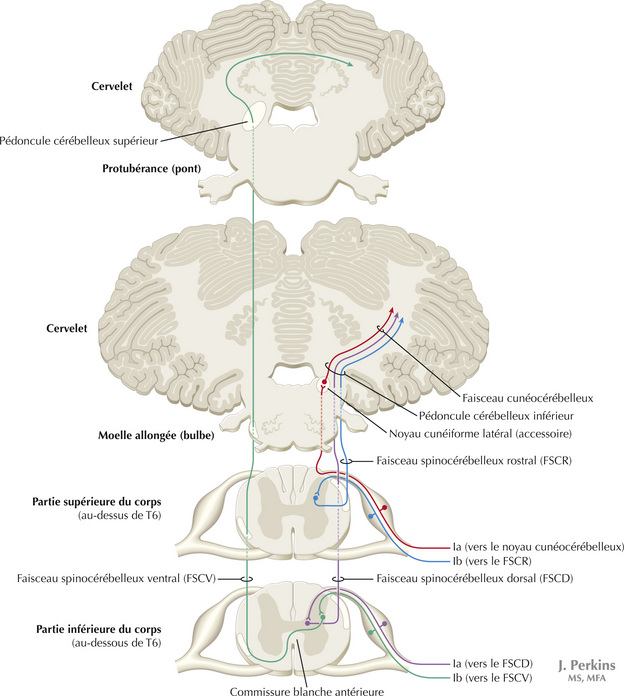
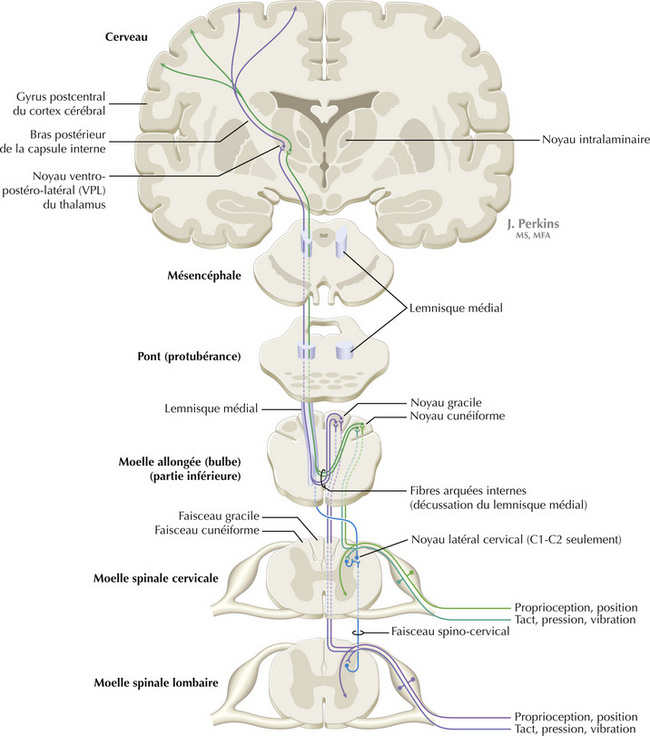
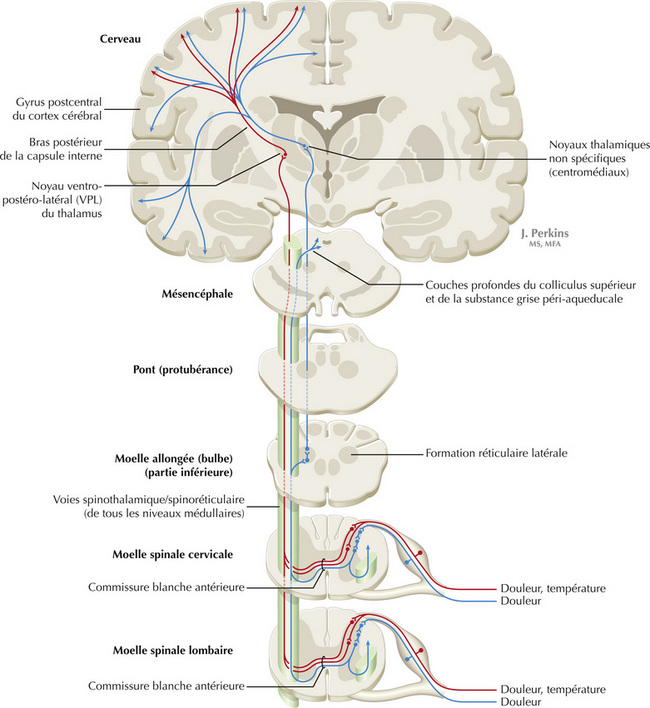
Figure 14.5 Système somatosensoriel : les voies spinothalamiques et spinoréticulaires et les modalités protopathiques
Les axones somatosensoriels amyéliniques (fibres C) et les petits axones myélinisés (fibres Aδ) qui véhiculent l’information nociceptive (rapide, douleur localisée), la sensation de température ainsi que le tact léger en déplacement, se projettent sur les neurones des couches I et V de la corne dorsale. Ceux-ci croisent la ligne médiane et forment la voie spinothalamique qui se projette sur le noyau ventro-postéro-latéral du thalamus (en rouge). Ce pool de neurones du noyau ventro-postérolatéral est différent du pool des neurones qui reçoivent les afférences des noyaux gracile et cunéiforme de la colonne dorsale. Ces neurones thalamiques se projettent à leur tour sur le cortex somatosensoriel secondaire (SII) et le cortex sensoriel primaire. Les fibres sensorielles primaires C se terminent aussi dans la corne dorsale et constituent un grand réseau en cascade, qui produit des projections bilatérales sur la voie spinoréticulaire (en bleu). Ce système se projette principalement sur la formation réticulaire d’où sont issues des projections polysynaptiques vers les noyaux non spécifiques du thalamus : médial, dorsal et antérieur. Certaines fibres spinoréticulaires se projettent sur les couches profondes du colliculus supérieur (voie spinotectale) et la substance grise péri-aqueducale. Les régions corticales cingulaire, insulaire et préfrontale traitent ensuite l’information de manière à interpréter l’information nociceptive liée aux douleurs lentes et fulgurantes.
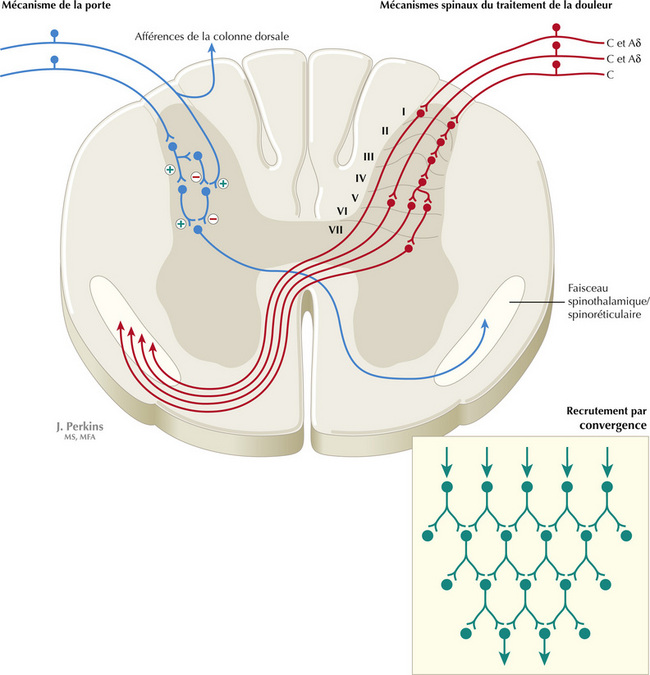
Figure 14.6 Traitement de la douleur par les voies spinothalamiques et spinoréticulaires de la moelle spinale






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


