15 Systèmes Moteurs
15.1 Motoneurones périphériques α et γ
15.2 Distribution des motoneurones périphériques dans la moelle spinale
15.3 Distribution des motoneurones périphériques dans le tronc cérébral
15.4 Voies efférentes corticales
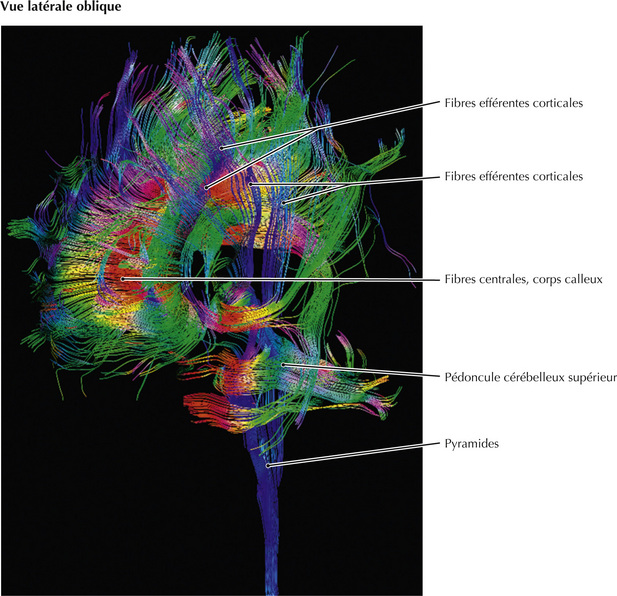
15.5 Imagerie en couleur des voies corticales efférentes
15.8 Projections corticospinales dans la moelle spinale
15.10 Faisceaux vestibulospinaux
15.11 Voies réticulospinale et corticoréticulaire
15.12 Faisceaux tectospinal et interstitiospinal
15.13 Terminaisons spinales des principales voies descendantes corticales motrices
15.14 Contrôle central des mouvements oculaires
15.15 Contrôle central de la respiration
15.16 Subdivisions fonctionnelles du cervelet
15.17 Réseau neuronal cérébelleux
15.18 Schéma des principales connexions afférentes du cervelet
15.19 Voies afférentes cérébelleuses
15.20 Voies cérébelleuses efférentes
15.21 Voies vestibulocérébelleuses et cérébellovestibulaires
15.22 Schéma des voies efférentes cérébelleuses destinées aux motoneurones centraux
Noyaux gris centraux (ganglions de la base)
15.23 Connexions des ganglions de la base
15.24 Principaux réseaux des ganglions de la base et neurotransmetteurs
15.25 Boucles fonctionnelles parallèles des ganglions de la base
Motoneurones périphériques
Les groupes de motoneurones périphériques contiennent à la fois les motoneurones a qui innervent les fibres musculaires squelettiques (fibres extrafusales) et les motoneurones y qui innervent les petits éléments contractiles des fuseaux neuromusculaires (fibres musculaires intrafusales), à l’exception du noyau du nerf facial, qui innerve les muscles faciaux. En effet, les muscles faciaux ne contiennent pas de fuseaux neuromusculaires et n’ont donc pas de motoneurones γ. Les motoneurones α régulent la contraction des muscles squelettiques afin de produire le mouvement. Les motoneurones γ régulent la sensibilité des fuseaux neuromusculaires qui, par leurs fibres afférentes la et Il, modulent l’excitabilité des motoneurones α
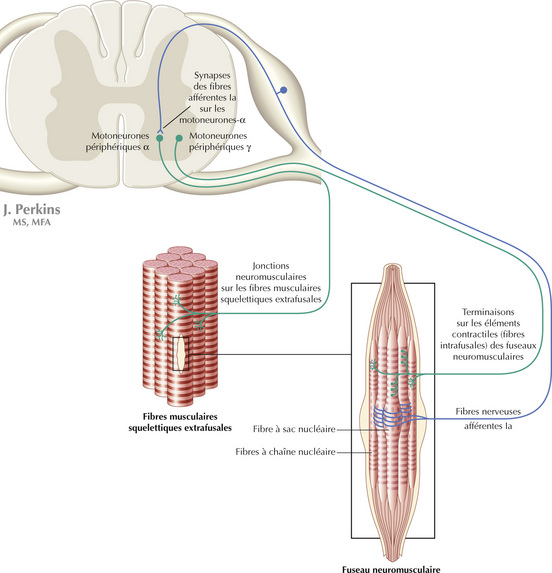
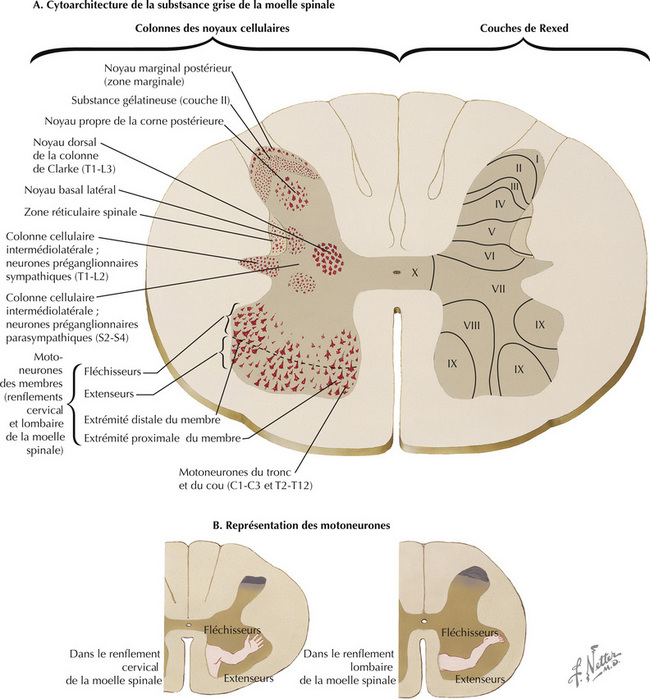
Les motoneurones périphériques sont regroupés en amas dans la corne antérieure (ventrale) de la moelle spinale, représentés dans la couche IX de Rexed. Ces groupes distincts sont chacun subordonnés à l’innervation de groupes musculaires squelettiques distincts et sont organisés de manière topographique. Les motoneurones périphériques destinés à la musculature du tronc et du cou sont en position médiane, tandis que ceux destinés aux muscles des extrémités distales sont en position latérale. À l’intérieur des segments de la moelle spinale, les motoneurones périphériques destinés aux fléchisseurs sont en position dorsale, tandis que ceux destinés aux extenseurs sont en position ventrale.
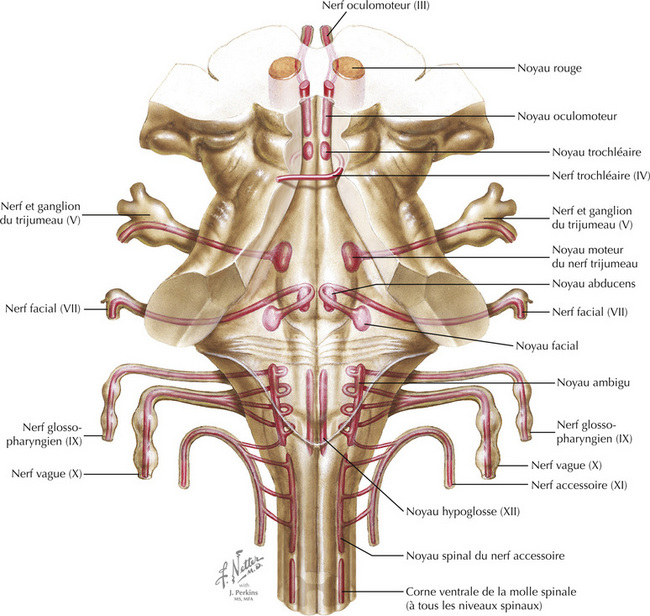
Les motoneurones périphériques sont situés dans des colonnes médiale et latérale d’une vue longitudinale du tronc cérébral. La colonne médiale (motoneurones périphériques des noyaux oculomo- teur, trochléaire, abducens et hypoglosse) provient de la voie efférente somatique générale ; la colonne latérale (motoneurones périphériques des noyaux moteur du V, facial, ambigu et spinal accessoire) provient de la voie efférente spécifique viscérale. Les motoneurones périphériques sont situés dans une colonne longitudinale à tous les niveaux de la corne ventrale (antérieure) de la moelle spinale.
Motoneurones centraux
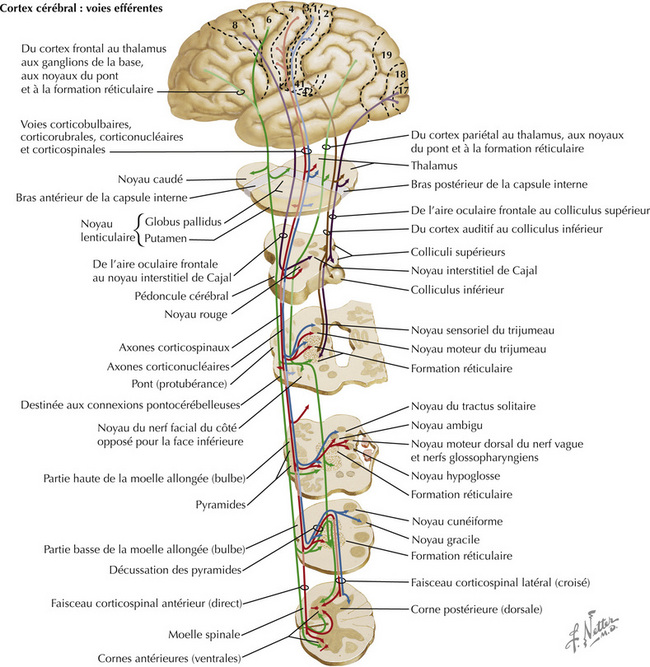
Les neurones du cortex moteur primaire (aire 4), du cortex prémoteur et de l’aire motrice supplémentaire (aire 6) envoient leurs axones aux ganglions de la base (noyau caudé et putamen), au thalamus (noyaux ventroantérieur et ventrolatéral), au noyau rouge, aux noyaux du pont, aux noyaux moteurs des nerfs crâniens des deux côtés, et à la corne ventrale de la moelle spinale, essentiellement au côté opposé. Ces axones forment la voie (ou faisceau ou tractus) corticospinale, la voie corticobulbaire, les projections corticostriées, corticopontiques, corti- cothalamiques et, enfin, les connexions corticales aux motoneurones centraux du tronc cérébral (aires motrices de la formation réticulaire, noyau rouge, colliculus supérieur). Les neurones du cortex sensoriel (aires 3, 1, 2) envoient leurs axones essentiellement aux noyaux sensoriels secondaires (fibres corticonucléaires) qui régulent l’information afférente de la voie lemniscale destinée à une interprétation consciente. Les neurones de l’aire oculaire frontale (aire 8) se projettent sur le colliculus supérieur, les centres de poursuite horizontale et verticale du tronc cérébral et le noyau interstitiel de Cajal, afin de coordonner les mouvements oculaires volontaires et les mouvements associés de la tête. D’autres régions du cortex sensoriel envoient leurs axones aux structures du thalamus et du tronc cérébral qui régulent l’information afférente sensorielle lemniscale. Certaines fibres efférentes corticales se projettent sur des structures limbiques comme les noyaux amygdaloïdes, la formation hippocampique et les noyaux du septum.
Cette image en tenseur de diffusion met en évidence les voies efférentes corticales selon un plan oblique latéral. Ces voies, représentées en bleu et provenant de zones très étendues du cortex cérébral, confluent vers le prosencéphale, le thalamus, le tronc cérébral, le cervelet et la moelle spinale. D’autres voies associatives corticales sont représentées en vert (direction antéro-postérieure) et les voies commissurales sont représentées en rouge (direction latérale, droite-gauche).
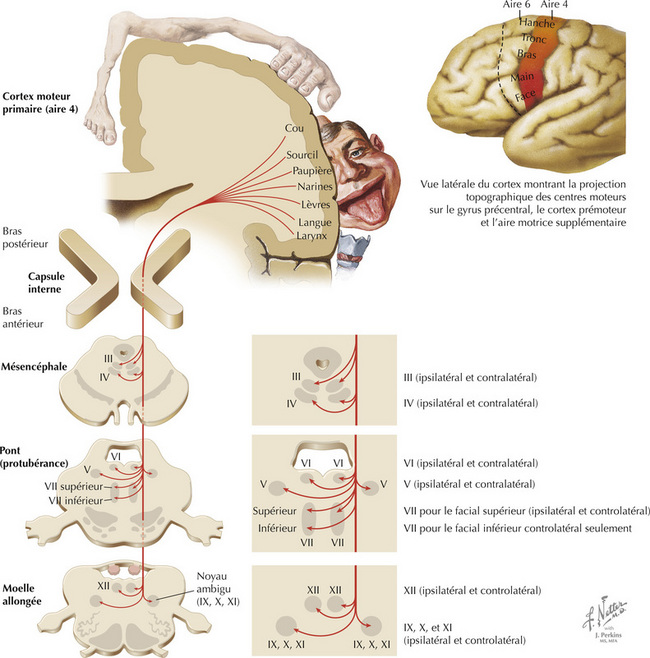
Le faisceau corticobulbaire naît principalement de la partie latérale du cortex moteur primaire (aire 4). Les axones qui en sont issus se projettent, par le genou de la capsule interne, dans le pédoncule cérébral, la base du pont et les pyramides bulbaires ipsilatérales. Les axones vont jusqu’aux noyaux moteurs des nerfs crâniens ipsi- et controlatéraux, sauf la partie du noyau moteur du nerf facial (VII) destinée aux muscles de l’expression dans la partie inférieure de la face, qui ne reçoivent que des projections controlatérales. Les projections corticobulbaires au noyau de l’hypoglosse sont essentiellement controlatérales. Celles qui atteignent le noyau du nerf spinal accessoire sont essentiellement ipsilatérales. Contrairement à la paralysie de Bell (atteinte du VII) où la totalité de l’hémiface ipsilatérale est paralysée, les lésions de la voie corticobulbaire ont pour conséquence un affaissement de la partie inférieure de l’hémiface controlatérale lors du mouvement volontaire (caractéristiques de la paralysie faciale d’origine centrale).
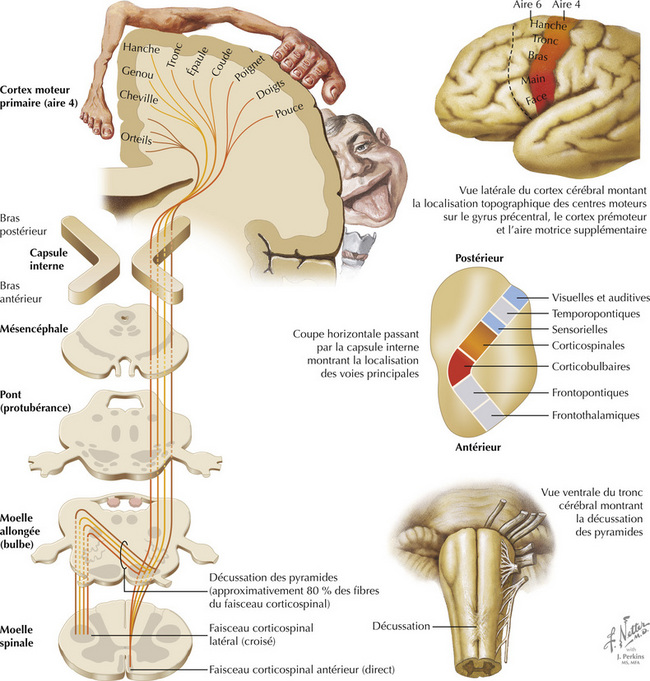
La partie motrice du faisceau corticospinal naît de neurones de taille variée, principalement dans le cortex moteur primaire (aire 4), le cortex prémoteur et l’aire motrice supplémentaire (aire 6). Le cortex sensoriel primaire (aires 3, 1, 2) apporte au faisceau corticospinal des axones qui se terminent essentiellement dans les noyaux sensoriels secondaires afin de réguler l’information afférente sensorielle lemnis- cale. Le faisceau corticospinal traverse le bras postérieur de la capsule interne, la région moyenne du pédoncule cérébral, de nombreux faisceaux d’axones de la base du pont et la pyramide bulbaire du même côté. La plupart de ces axones — en moyenne 80 %, mais ceci est variable entre les individus —, croisent la ligne médiane au niveau de la jonction bulbospinale : c’est la décussation des pyramides. Ils descendent dans le faisceau corticospinal latéral, dans le funiculus latéral de la moelle spinale et font synapse sur les motoneurones α et les motoneurones γ à la fois directement et indirectement par le biais d’interneurones. Les axones du faisceau corticospinal qui ne décus- sent pas descendent dans le faisceau corticospinal antérieur, dans le funiculus antérieur de la molle spinale, puis décussent au niveau approprié par la commissure blanche antérieure, pour se terminer directement et indirectement sur les motoneurones controlatéraux à leurs neurones corticaux d’origine. Seule une très petite proportion des axones du faisceau corticospinal se terminent directement sur les motoneurones ipsilatéraux de la molle spinale.
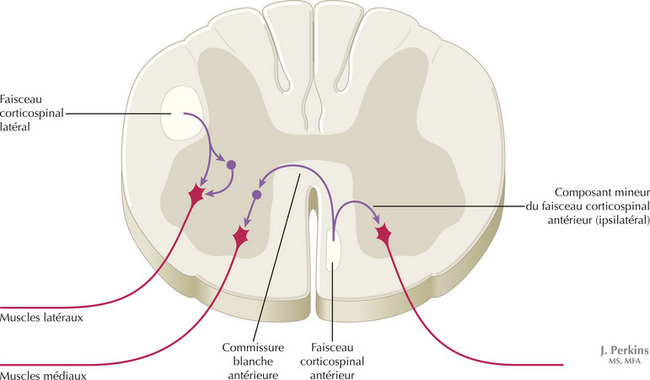
Les axones du faisceau corticospinal décussés sont associés aux axones du faisceau rubrospinal dans le funiculus latéral, ou faisceau pyramidal croisé ou faisceau pyramidal I atéral. La plupart de ces axones se terminent directement ou indirectement sur les motoneurones de la musculature distale commandant l’habileté des mains et des doigts. Les axones non croisés du faisceau corticospinal direct ou antérieur décussent principalement au niveau de la commissure blanche antérieure et se terminent directement et indirectement sur les motoneurones de la musculature médiane. Un petit nombre d’axones du faisceau corticospinal direct se projettent de manière ipsilatérale aux neurones centraux. Une lésion isolée du faisceau corticospinal dans une pyramide bulbaire a pour conséquence un déficit de la motricité fine de la main et des doigts controlatéraux. Toutes les autres lésions impliquant le faisceau corticospinal au niveau de la capsule interne, du pédoncule cérébral et du pont sont responsables à long terme d’une hémiplégie spastique controlatérale associant une hypertonie, une hyperréflexie et une réponse cutanée plantaire en extension. Une lésion du faisceau corticospinal latéral, dans la moelle spinale, a les mêmes conséquences, du même côté et au-dessous de la lésion.
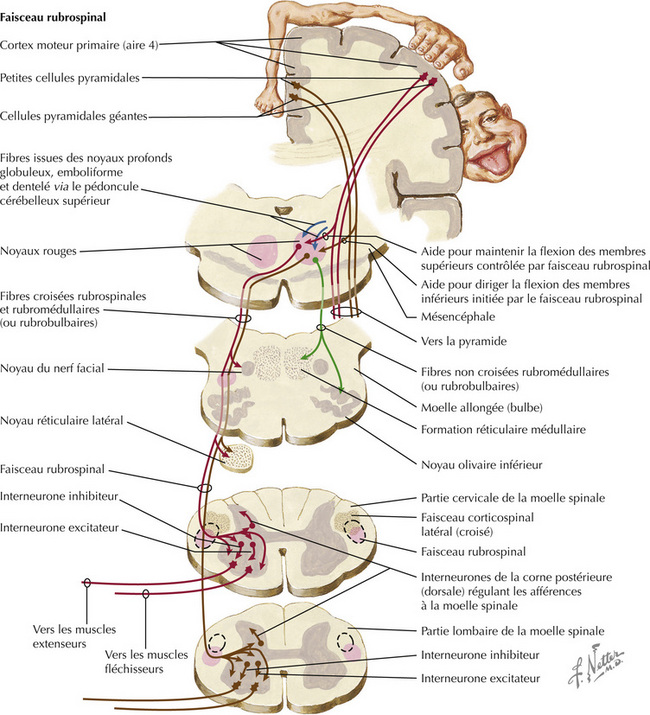
Le système cortico-rubro-spinal fonctionne comme un système corticospinal indirect qui régule les motoneurones périphériques de la moelle spinale. Le noyau rouge dans le mésencéphale reçoit des afférences parfaitement organisées sur le plan topographique du cortex moteur primaire (aire 4) ipsilatéral. Les axones du faisceau rubrospinal décussent au niveau de la décussation tegmentale ventrale et descendent dans le tronc cérébral latéral et le funiculus latéral de la moelle spinale, où ils sont mélangés aux axones du faisceau corticospinal latéral. Le faisceau rubrospinal se termine directement et indirectement sur les motoneurones a et les motoneurones g de la moelle spinale, en particulier ceux responsables des mouvements de flexion des extrémités. Le faisceau rubrospinal aide au contrôle des fléchisseurs des membres supérieurs et des membres inférieurs. Les lésions du faisceau rubrospinal dans la moelle spinale sont généralement concomitantes d’atteintes du faisceau corticospinal, tout comme les lésions corticorubrales au niveau de la capsule interne et du pédoncule cérébral. Ces lésions ont pour conséquence à long terme une hémiplégie spastique controlatérale. Les lésions du tronc cérébral caudales au noyau rouge ont pour conséquence la décérébration (spasticité en extension), reflétant ainsi la levée du contrôle des fléchisseurs de la voie rubrospinale des motoneurones des membres supérieurs. (Physiopathologie : cf. p. 386)






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


