1 Neurones
1.2 Les différents types de synapses
1.3 Les différents types de neurones
1.4 Les différents types de cellules gliales
1.5 La barrière hématoencéphalique
1.6 Myélinisation du système nerveux central et périphérique
1.7 Développement des gaines myéliniques des axones
1.8 Agrandissement d’une gaine de myéline dans le système nerveux central
1.9 Les différents types de synapses chimiques
1.10 Morphologie de la synapse
1.11 Potentiel de repos du neurone
1.12 Potentiels gradués des neurones
1.14 Propagation du potentiel d’action
1.16 Classification des fibres nerveuses périphériques par leur diamètre et leur vitesse de conduction
1.17 Électromyographie et études de vitesses de conduction
1.18 Inhibition présynaptique et post-synaptique
1.19 Sommation spatiale et temporelle
1.20 Profils normaux de décharge électrique des neurones corticaux et origine et propagation des crises d’épilepsie
Structure
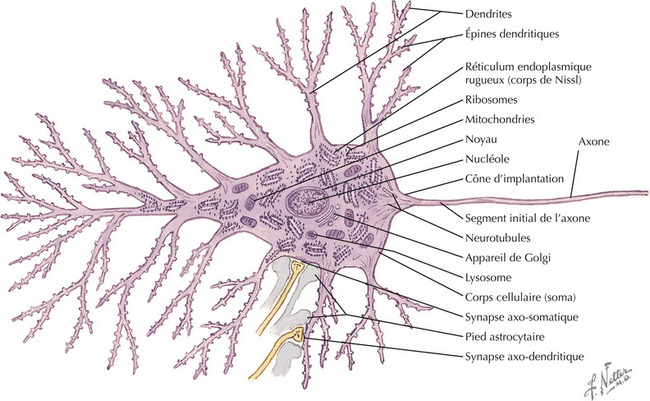
La structure du neurone reflète ses caractéristiques fonctionnelles. L’information afférente parvient principalement au neurone par les terminaisons axonales qui se projettent sur le corps cellulaire et les dendrites. Ces synapses sont isolées et protégées par les pieds astrocy- taires. Les dendrites constituent habituellement la majeure partie de la surface du neurone. Les épines dendritiques, qui sont des protrusions situées sur les branches dendritiques, sont les cibles spécifiques des synapses axo-dendritiques. Chaque type neuronal a sa propre arborisation dendritique, avec ses caractéristiques spécifiques. La taille du corps cellulaire du neurone est très variable, de quelques microns à plus de 100 |im de diamètre. Le cytoplasme du neurone contient un abondant réticulum endoplasmique rugueux (RER), qui témoigne d’une intense activité de synthèse protéique pour la survie et le fonctionnement de la cellule. L’appareil de Golgi assure le conditionnement des molécules impliquées dans la transmission du signal en vue de leur transport et de leur libération. Les besoins énergétiques énormes des neurones nécessitent un grand nombre de mitochondries, en particulier pour le fonctionnement des pompes ioniques et le maintien du potentiel de membrane. Chaque neurone a habituellement un seul axone (il peut n’en avoir aucun). Le corps cellulaire s’effile au niveau du cône d’implantation, pour se continuer par le segment initial de l’axone, qui contient les canaux Na+ et constitue le site d’initiation du potentiel d’action. La longueur de l’axone est variable et peut dépasser 1 mètre. Les axones de plus de 1 à 2 |im de diamètre sont isolés par les feuillets myéliniques ; ces feuillets sont formés par les cellules oligodendrogliales dans le système nerveux central (SNC) et par les cellules de Schwann dans le système nerveux périphérique (SNP). Un axone peut avoir jusqu’à 500 000 terminaisons, se projetant sur un site très localisé (par exemple, les projections somatosensorielles du tact fin discriminatif) ou bien se distribuant dans des régions très différentes du cerveau (par exemple, les projections axonales noradrénergi- ques du locus cœruleus). Les axones des grands neurones (neurones de Golgi de type I) se projettent loin du corps cellulaire et de l’arbre dendritique, ceux des petits neurones (neurones de Golgi de type II, neurones de réseau local, interneurones) se projettent localement, près du corps cellulaire et de l’arbre dendritique. Chaque type de neurone étant hautement spécialisé, il n’y a pas de neurone « représentatif »», bien que les cellules pyramidales ou les motoneurones de deuxième ordre soient souvent décrites comme modèle de neurone.
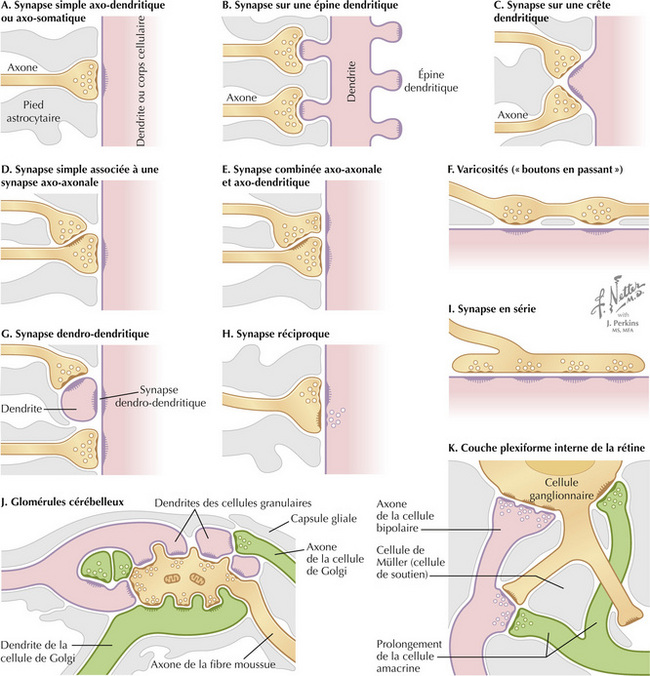
La synapse est le lieu où le potentiel d’action qui arrive déclenche la libération d’un ou plusieurs neurotransmetteurs dans la fente synaptique (habituellement large de 20 nm). Cette libération de neurotransmetteurs se fait par un couplage excitation-sécrétion impliquant un flux entrant calcique. Le neurotransmetteur se fixe sur son récepteur situé sur la membrane cible post-synaptique, dont il modifie le potentiel par rapport à l’état de repos. Ces potentiels post-synaptiques sont dénommés potentiels gradués (ou locaux). La plupart de ces synapses sont des synapses axo-dendritiques ou axo-somatiques. Des synapses spécialisées, comme les synapses réciproques, ou des ensembles complexes de synapses en interaction, jouent un rôle régulateur spécifique sur l’excitabilité des neurones cibles. Les synapses dendro-dendritiques interviennent dans la décharge coordonnée de mêmes groupes neuronaux (comme les neurones du noyau phrénique permettant la contraction du muscle diaphragmatique).
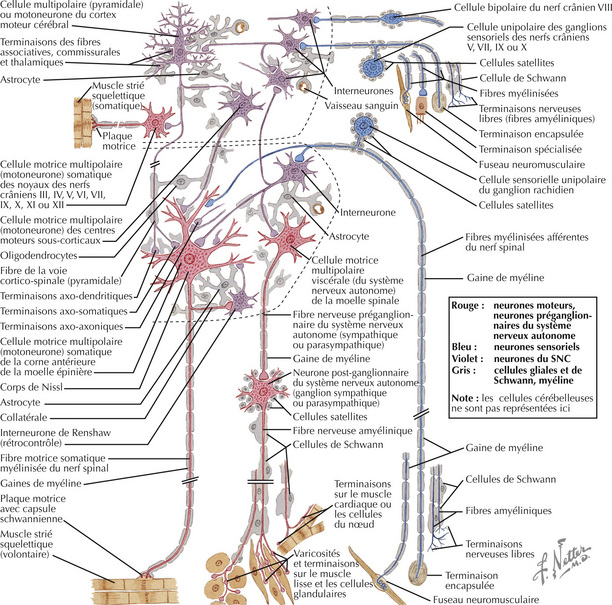
Les interneurones locaux et les neurones de projection peuvent être caractérisés par leur taille, leur arborisation dendritique et leurs projections axonales. Dans le SNC (délimité dans la figure par les pointillés), les cellules gliales (astrocytes, microglie, oligodendro- glie) protègent les neurones et subviennent à leurs besoins. Les cellules de Schwann et les cellules satellites jouent le même rôle dans le SNP. Les neurones sensoriels primaires (en bleu) conduisent l’information afférente sensorielle : les stimuli sont traduits sous forme de signaux électriques transportés jusqu’au SNC. L’information efférente du SNC (en rouge) est soit motrice jusqu’aux fibres musculaires striées squelettiques via la jonction neuromusculaire, soit appartient au système nerveux autonome et conduit l’information jusqu’au ganglion autonome via les fibres préganglionnaires. Du ganglion partent les fibres post-ganglionnaires qui ont pour cible le muscle cardiaque, le muscle lisse, les glandes sécrétoires, les cellules métaboliques ou encore les cellules du système immunitaire. Les corps cellulaires des motoneurones de 2e ordre et des fibres préganglionnaires du système nerveux autonome sont situés dans la moelle spinale (zone pointillée inférieure) ou bien dans le cerveau (zone pointillée supérieure).
Les astrocytes isolent les neurones du SNC et leurs synapses. Ils tamponnent les ions K+ et ont un rôle trophique en favorisant la croissance du neurone et le signal cellulaire. Les cellules oligoden- drogliales myélinisent les axones du SNC. Les cellules microgliales sont des cellules « éboueurs » qui participent à la phagocytose, aux réponses inflammatoires, à la sécrétion de cytokines et de facteurs de croissance et, dans une certaine mesure, à l’activité immunitaire du SNC. Les cellules périvasculaires, comme les péricytes par exemple, ont des activités analogues au voisinage des vaisseaux sanguins. Les cellules de Schwann myélinisent les neurones périphériques et jouent un rôle dans la nutrition, la croissance et la réparation de ces neurones. Des lymphocytes T activés peuvent pénétrer dans le SNC et le parcourir pendant environ 24 heures pour assurer sa surveillance immunitaire.
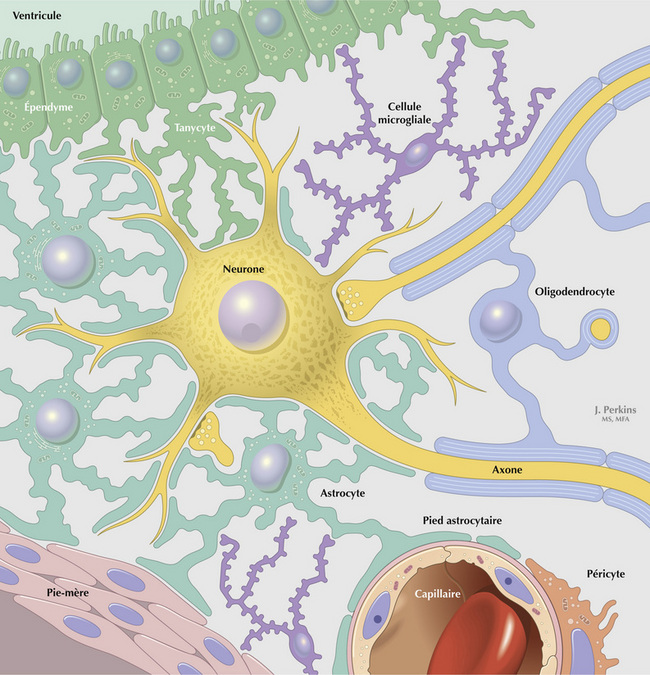
La barrière hématoencéphalique (BHE) est constituée par l’interface cellulaire entre le sang et le SNC. Son rôle est d’empêcher l’intrusion de grosses molécules potentiellement neurotoxiques et d’assurer l’homéostasie locale afin de favoriser le fonctionnement optimal du neurone et de la cellule gliale. L’élément clé de la BHE est constitué par la cellule endothéliale du capillaire. Ces cellules endothéliales sont unies les unes aux autres par des jonctions serrées, ce qui limite l’accès des grosses molécules (des médicaments en particulier) au SNC. Elles ont une activité pinocytique réduite mais possèdent des systèmes de transport sélectifs et spécifiques pour les substrats essentiels à la production énergétique au métabolisme des acides aminés dans le SNC. Les pieds astrocytaires entourent les lames basales des cellules endothéliales, permettant le transfert de métabolites du sang vers les neurones et, éventuellement, la régulation de l’expression endothéliale de gènes spécifiques. Ils permettent également de tamponner le K+ et les neurotransmetteurs en excès de l’espace interstitiel.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


