Chapitre 1 Développement embryologique du membre supérieur
APPARITION DES BOURGEONS DES MEMBRES
Chez l’Homme, les bourgeons des membres apparaissent entre les troisième et quatrième semaines du développement embryonnaire. Les membres supérieurs précèdent les membres inférieurs d’environ quatre jours. Leurs schémas de formation reposent sur des signaux moléculaires et des séquences cellulaires de chronologie similaire [4].
Ainsi, au 24e jour apparaissent des protubérances sur les flancs, dans la région cervicale de l’embryon. Ils sont constitués d’un contingent de cellules provenant du mésoderme latéral (ou somatopleure), qui sera à l’origine de l’ensemble des éléments squelettiques (figure 1.1a et b). Ce bourgeon est recouvert par l’ectoderme dont l’épithélium se condense et se stratifie précocement à son extrémité distale pour former la crête ectodermique apicale (CEA). Elle représente une des structures clés de la régulation de la croissance du bourgeon du membre (figure 1.1c).
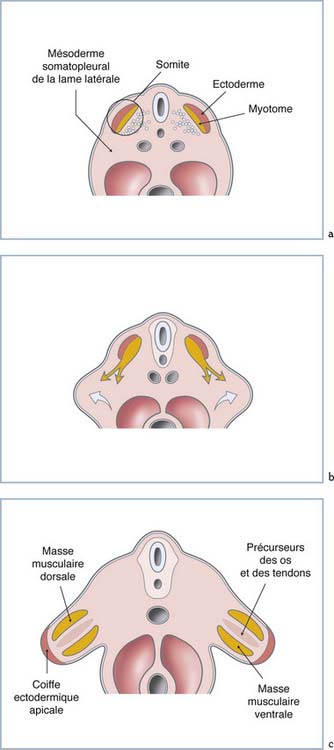
L’apparition des bourgeons semble être initiée par des signaux moléculaires faisant intervenir des membres de la famille des facteurs de croissance fibroblastiques (FGF : Fibroblast Growth Factor) [2]. Le FGF-8 en particulier semble jouer un rôle précoce dans cette initiation. En effet, la présence d’ARN messager de cette molécule est identifiée dans l’ectoderme de la zone présomptive du bourgeon de membre, avant même son apparition histologique. Ce facteur est initialement détecté au niveau du mésoderme intermédiaire (entre la colonne somitique et le mésoderme latéral) de la totalité des flancs de l’embryon. Son expression se limite rapidement au niveau du futur site d’apparition des bourgeons des membres supérieurs et inférieurs. Par ailleurs, l’implantation expérimentale de billes imprégnées de FGF-8 au niveau du flanc, entre les bourgeons supérieurs et inférieurs, initie la formation d’un membre surnuméraire. Ainsi, il apparaît que cette molécule peut initier un programme morphogénétique qui s’autonomisera jusqu’à assurer la formation complète d’un membre. De même, les FGF-4 et FGF-7 sont capables d’induire expérimentalement toutes les étapes de la formation de membres ectopiques, mais ces molécules ne sont pas exprimées dans ces sites lors des phases physiologiques de l’initiation du bourgeon des membres.
ALLONGEMENT DU BOURGEON DES MEMBRES
Dès son apparition sur les flancs de l’embryon, le bourgeon s’allonge selon une polarité proximo-distale, du fait de l’activité proliférative de son extrémité appelée « zone de progrès ». En effet, les cellules mésenchymateuses la constituant présentent une activité mitotique intense, régulée par des facteurs de croissance émis par la CEA à son contact. Celle-ci en module l’activité par l’émission concomitante de facteurs moléculaires tels que FGF-4 et FGF-8 la stimulant et d’autres, au contraire, tels que BMP-2 et BMP-4, qui inhibent la prolifération cellulaire [6]. L’exérèse expérimentale de la CEA aboutit à la formation d’un membre tronqué, d’autant plus proximalement que l’exérèse était précoce, avec absence complète des structures distales. Ce déficit transversal est réversible par un apport local de FGF-4 qui rétablit l’activité mitotique du front de prolifération. De plus, dans les conditions spécifiques de positionnement adéquat, ce facteur moléculaire rétablit la formation des structures anatomiques distales rendant compte de son rôle dans l’établissement d’informations positionnelles au sein du bourgeon de membre.
MORPHOGENÈSE
La morphogenèse des membres supérieurs se déroule de la 4e à la 8e semaine de développement. C’est au cours de ces étapes que se met rapidement en place leur anatomie (figure 1.2).

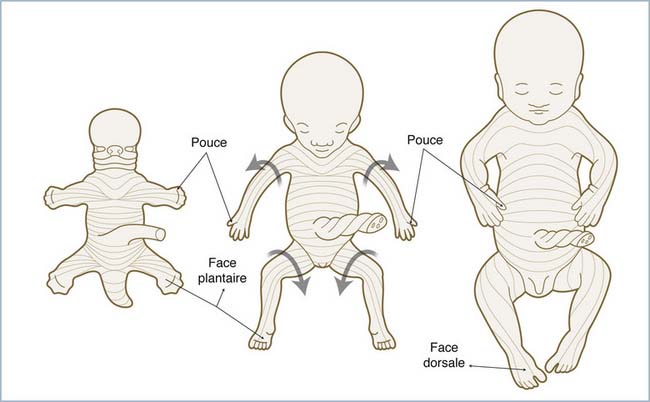
Au 47e jour, la racine du bourgeon du membre subit une flexion horizontale qui le réoriente d’une position initiale frontale vers un plan para-sagittal. Les membres subissent alors un mécanisme de torsion externe accompagnant la migration caudale de la ceinture scapulaire, de sorte que la face ventrale initiale du bourgeon devient sa face crâniale. Le coude qui se fléchit progressivement fait alors saillie en direction caudale. Cette rotation, moins marquée qu’au niveau des membres inférieurs, a pour conséquence de tordre, en spirale, le plan de segmentation de l’innervation sensitive (figure 1.3).






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


