Chapitre 2 Cartilage de croissance et périoste
Physe
ANATOMIE
Les os longs du squelette appendiculaire présentent une physe à chacune de leurs extrémités. Elles contribuent, de manière inégale, au gain de longueur de l’os en croissance [2]. Au niveau des membres inférieurs, la productivité dominante est située près du genou. Le fémur distal et les fibula et tibia proximaux contribuent en moyenne à 80 % de l’accroissement en longueur du membre. De même, pour les membres supérieurs, la productivité dominante est située loin du coude (humérus proximal, radius et ulna distaux). Les physes assurent leurs fonctions jusqu’en fin de puberté où la maturité squelettique s’accompagne de leur disparition définitive et signe la fin de la croissance. Cette disparition, asynchrone selon les localisations anatomiques, permet d’évaluer « l’âge osseux » de l’adolescent.
Contrairement au cartilage hyalin articulaire, le cartilage de conjugaison est le siège d’une activité métabolique intense. Celle-ci repose sur une vascularisation particulièrement riche provenant des cercles anastomotiques péri-articulaires [2]. Elle trouve son origine au niveau de trois systèmes distincts. Les vaisseaux épiphysaires contribuent à la vascularisation du versant épiphysaire du cartilage de croissance : les zones germinales et prolifératives. Les vaisseaux métaphysaires osseux, ainsi que les branches terminales des artères nourricières, assurent les échanges métaboliques de son versant métaphysaire : la zone de minéralisation et d’ossification. Un troisième réseau provient de la virole périchondrale et contribue à la vascularisation de la couronne périphérique du cartilage de conjugaison.
HISTOLOGIE
La physe est organisée en strates cellulaires successives composées de chondrocytes de stade de maturité croissante selon une polarité dirigée de l’épiphyse vers la métaphyse [6].
Son unité fonctionnelle peut être considérée comme une colonne dont les chondrocytes passent d’un stade de prolifération à celui d’hypertrophie, puis de minéralisation de la matrice extra-cellulaire, avant d’entrer en apoptose (mort cellulaire programmée) (figure 2.1). Ces étapes reproduisent celles de l’ostéogenèse enchondrale dans une distribution spatio-temporelle spécifique.
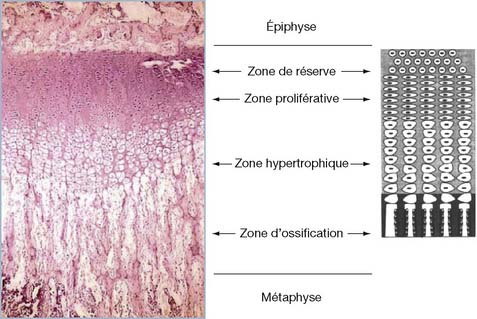
La première strate est constituée par les « cellules germinales ». Ces chondrocytes se distribuent, de manière désordonnée, dans une matrice extra-cellulaire abondante vascularisée par le réseau capillaire épiphysaire. Leur contenu cytoplasmique, riche en vacuoles lipidiques, suggère leur contribution métabolique à l’activité du cartilage. Ces cellules ne participent pas directement à l’accroissement en longueur de l’os. En effet, l’activité mitotique de cette première strate est très faible. Cependant, par sa croissance interstitielle, elle permet la migration centrifuge des chondrocytes en direction de l’encoche d’ossification de Ranvier. Ce processus, actif jusqu’à la maturité squelettique, contribue à l’expansion diamétrale de la physe [5].
PHYSIOLOGIE
Régulation biologique
Les facteurs de croissance locaux sont exprimés par les chondrocytes eux-mêmes et agissent selon des mécanismes autocrines, c’est-à-dire sur les cellules mêmes qui les ont sécrétés, ainsi que par des mécanismes paracrines, s’exerçant sur celles de leur environnement. Ils contribuent à la régulation intrinsèque de la production de la physe [3]. Ihh (Indian Hedgehog) et PTHrP (Parathormone Related Peptide) équilibrent la maturation de ces chondrocytes. En particulier, Ihh stimule la synthèse de la matrice extra-cellulaire, la prolifération ainsi que la maturation des chondrocytes et leur passage irréversible de cellules à potentiel prolifératif au stade de chondrocytes hypertrophiques. Le PTHrP s’oppose à cette différenciation, les maintenant dans le pool des cellules prolifératives. Les BMP (Bone Morphogenetic Protein) stimulent l’expression de Ihh. Au contraire, le FGF (Fibroblast Growth Factor) exerce des effets antagonistes en l’inhibant.
Régulation mécanique
Les forces s’opposant à la croissance ne sont pas connues avec précision. Le manchon de périoste qui s’insère sur les épiphyses, aux extrémités opposées des os longs, exerce une contrainte statique, d’amplitude méconnue, au niveau des deux physes qu’il « ponte » (figure 2.2). Cependant, son effet freinateur sur la croissance a été mis en évidence par diverses expérimentations. En effet, la libération du « frein périostique », par sa section transversale ou son excision, débride la croissance de la physe qui subit une stimulation transitoire.
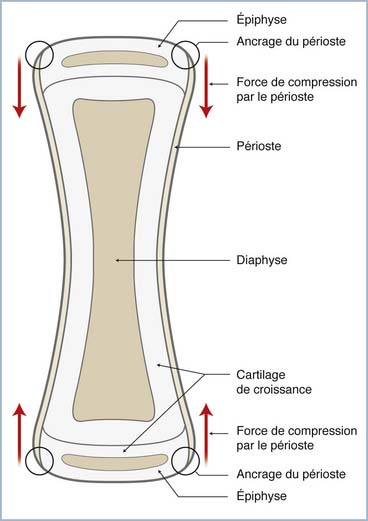
Au contraire, la distraction maintenue à une faible vitesse, limitant les déformations sous un seuil de rupture, entraîne une accélération de la croissance par hyperplasie des couches prolifératives : c’est la chondrodiastase [3]. Pour des vitesses élevées, entraînant des déformations supérieures au seuil à la rupture, des fractures-séparations ou épiphysiolyses se produisent, et un allongement progressif peut être obtenu selon les mécanismes biologiques équivalant à ceux de l’ostéogenèse par distraction observés après ostéotomie.
PHYSE EN SITUATION PATHOLOGIQUE
Une structure vulnérable
Sur le plan mécanique, les physes représentent des zones « transitionnelles » cartilagineuses interposées entre deux segments osseux plus rigides. Elles constituent des zones de concentration des sollicitations mécaniques, responsables de l’extrême fréquence des lésions auxquelles elles sont exposées dans la pathologie traumatique [4–7] (figure 2.3) ainsi que lors des sollicitations répétées liées à l’activité sportive intensive.
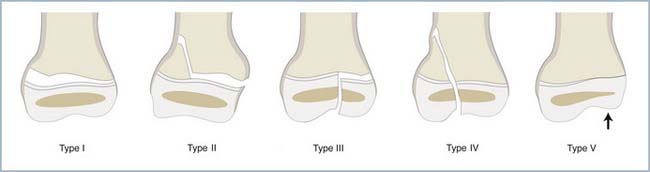
2.3 Classification de Salter et Harris des fractures épiphysaires en cinq types [7]. Les types I et II passent dans l’épaisseur de la physe, entre la couche sériée et hypertrophique : la couche germinale reste intacte, et le risque d’épiphysiodèse est peu élevé. À l’inverse, les types III et IV traversent complètement le cartilage de croissance, risquant de léser la zone germinative, et donc d’entraîner une épiphysiodèse définitive. Le type V a la particularité de ne pas être forcément visible d’emblée à la radiographie, mais il est dû à une impaction sévère de la physe, avec un risque de destruction complète.
Related posts:
 13: Ostéotomies supra-condyliennes de varisation et valgisation
13: Ostéotomies supra-condyliennes de varisation et valgisation
 11: Ostéotomie de dérotation humérale pour séquelle de paralysie obstétricale du plexus brachial
11: Ostéotomie de dérotation humérale pour séquelle de paralysie obstétricale du plexus brachial
 23: Exostoses sous-unguéales
23: Exostoses sous-unguéales
 16: Fractures récentes et anciennes de Monteggia
16: Fractures récentes et anciennes de Monteggia
 28: Duplications du pouce de types 1, 2 et 4
28: Duplications du pouce de types 1, 2 et 4
 31: BrÛlures des mains
31: BrÛlures des mains
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


