Chapitre 9
Les muscles lombaux et leurs fascias
Le rachis lombal est entouré de muscles qui peuvent être divisés en trois groupes pour des raisons fonctionnelles et dans un but descriptif :
• le grand psoas recouvrant la partie antérolatérale du rachis lombal ;
• les intertransversaires latéraux et le carré de lombes qui relient et recouvrent les processus transverses antérieurement ;
• les muscles lombaux postérieurs qui reposent en arrière et recouvrent les éléments postérieurs du rachis lombal.
GRAND PSOAS
Le grand psoas est un long muscle qui émerge de la face antéro-externe du rachis lombal et descend au-dessus de la bordure du pelvis pour s’insérer sur le petit trochanter du fémur. C’est essentiellement un muscle de la cuisse dont l’action principale est la flexion de la hanche.
Le muscle grand psoas possède des insertions sur le rachis lombal qui sont diverses mais précises (figure 9.1). À chaque niveau segmentaire de T12–L1 à L4–L5, il s’insère sur les trois quarts internes environ de la surface antérieure du processus transverse, sur le disque intervertébral et sur la bordure des corps vertébraux adjacente au disque [1]. Un faisceau supplémentaire émerge du corps vertébral de L5. Le muscle est censé émerger généralement d’une voûte tendineuse recouvrant la face latérale du corps vertébral [2]. Cependant, une dissection minutieuse [1] révèle que ces voûtes ne constituent rien d’autre que la partie interne du fascia musculaire profond et que celui-ci n’offre aucune origine particulière supplémentaire. Les fibres musculaires les plus internes contournent le muscle et sont ancrées directement sur le bord supérieur du corps vertébral. Le fascia forme cependant une arcade au fond du psoas, sur la surface externe du corps vertébral, laissant un espace entre l’arche et l’os transmettant les artères lombales et les veines (voir chapitre 11).
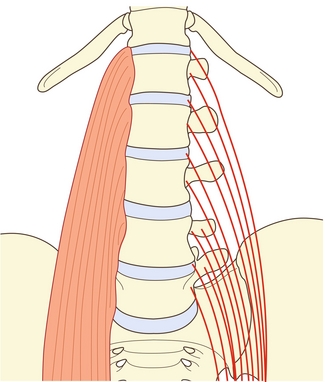
Figure 9.1 Psoas (P). À chaque niveau segmentaire, le grand psoas s’attache au processus transverse, au disque intervertébral et aux bords de la vertèbre adjacente.
Les fibres musculaires du disque intervertébral L4–L5, le corps de L5 et le processus transverse de L5 forment le faisceau de fibres intramusculaires le plus profond et le plus inférieur. Ces fibres sont systématiquement recouvertes par des fibres provenant du disque, des bords vertébraux et du processus transverse, sur des niveaux de plus en plus élevés. Sur une coupe transversale, le muscle est disposé en couches circonférentielles ; les fibres des étages supérieurs forment la surface externe du muscle et celles des étages inférieurs recouvrent séquentiellement en profondeur sa substance. À l’intérieur du muscle, les faisceaux des segments lombaux individuels ont la même longueur, de telle sorte que ceux de L1 deviennent tendineux avant ceux des vertèbres inférieures successives. Cette morphologie isométrique indique que le muscle est conçu exclusivement pour agir sur la hanche [1].
Les études biomécaniques révèlent que le grand psoas n’a qu’une faible action sur le rachis lombal en flexion et en extension. Ses fibres sont disposées pour permettre l’extension des segments lombaux supérieurs et la flexion des segments lombaux inférieurs. Toutefois, les fibres agissent très près des axes de rotation des vertèbres lombales et ne peuvent exercer que de très petits moments, même en contraction maximale [1]. Cela est en contradiction avec la possibilité d’une action significative du grand psoas sur le rachis lombal. Il utilise plutôt le rachis lombal comme base à partir de laquelle il agit sur la hanche.
Cependant, le grand psoas exerce potentiellement une énorme charge compressive sur les disques lombaux inférieurs. La proximité de la ligne d’action du muscle avec les axes de rotation minimise sa capacité de flexion, mais maximise la compression axiale exercée. En contraction maximale, dans une activité comme les sit-ups (abdominaux), les deux psoas peuvent exercer une charge compressive sur le disque L5–S1 équivalente à une charge d’environ 100 kg [1].
INTERTRANSVERSAIRES LATÉRAUX
Les intertransversaires latéraux sont constitués de deux parties : les intertransversaires latéraux ventraux et les intertransversaires latéraux dorsaux. Les intertransversaires ventraux relient les bords des processus transverses successifs, alors que les dorsaux unissent chacun un tubercule accessoire au processus transverse inférieur (figure 9.2). Les intertransversaires ventraux et dorsaux sont tous les deux innervés par les rameaux ventraux des nerfs spinaux lombaux [3] et ne peuvent donc pas être classés parmi les muscles rachidiens, tous innervés par les rameaux dorsaux (voir chapitre 10). Du fait de leurs insertions et de leur innervation, les intertransversaires ventraux et dorsaux sont considérés équivalents aux muscles intercostaux et aux élévateurs des côtes de la région thoracique [3].
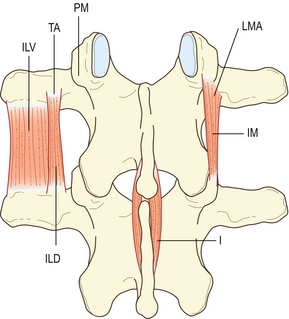
Figure 9.2 Muscles intersegmentaires courts. I : interépineux ; ILD : intertransversaires latéraux dorsaux ; ILV : intertransversaires latéraux ventraux ; IM : intertransversaires médiaux ; LMA : ligament mamillaire accessoire ; PM : processus mamillaire ; TA : tubercule accessoire.
CARRÉ DES LOMBES
Le carré des lombes est un muscle large, plus ou moins rectangulaire qui recouvre à peu près les deux tiers externes de la surface antérieure des processus transverses de L1 à L4 et se prolonge latéralement au-delà des extrémités des processus transverses, sur quelques centimètres. En le détaillant, le muscle est un agrégat complexe de multiples fibres dirigées obliquement et longitudinalement, reliant les processus transverses lombaux, l’ilium et la 12e côte [4] (figure 9.3).
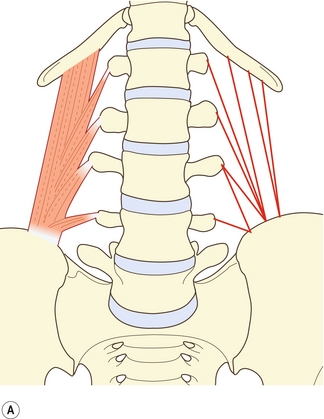
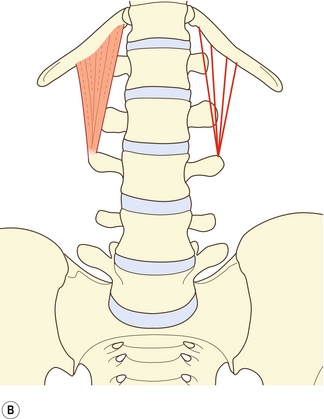
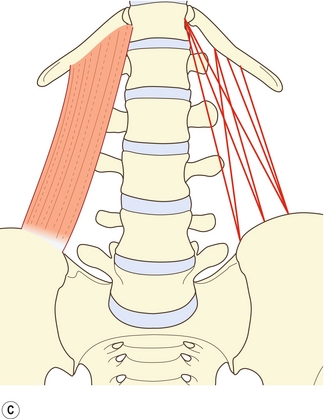
Figure 9.3 Couches et faisceaux les plus courants du carré des lombes. A : Couche postérieure. B : Couche intermédiaire ; C : Couche antérieure.
On peut considérer que ce muscle est composé de quatre types de faisceaux disposés en trois couches [5]. Les fibres iliocostales relient l’ilium à la 12e côte. Les fibres iliolombales relient l’ilium aux processus transverses lombaux. Un quatrième type de faisceaux relie l’ilium au corps de la 12e vertèbre thoracique. Quelques faisceaux relient parfois les processus transverses lombaux au corps de la 12e vertèbre thoracique.
La couche postérieure (figure 9.3A) est composée inférieurement et médialement des faisceaux iliolombaux, et latéralement des faisceaux iliocostaux [5]. Les fibres iliolombales émergent de la crête iliaque et s’insèrent plus constamment sur les processus transverses des trois lombales supérieures. Quelques faisceaux s’insèrent aussi parfois sur le processus transverse de L4.
La couche intermédiaire (figure 9.3B) émerge typiquement de la surface antérieure du processus transverse de L3 par un tendon commun. Ces faisceaux irradient vers la face inférieure et antérieure de la moitié médiale environ de la 12e côte [5]. Ces faisceaux sont parfois reliés à ceux des processus transverses de L2, L4 et L5.
La couche antérieure (figure 9.3C) est composée de fibres plus ou moins parallèles provenant de la crête iliaque et se dirigeant vers le haut. Les fibres les plus latérales s’insèrent sur la face antéro-inférieure de la 12e côte. Les fibres les plus médiales s’insèrent sur un tubercule de la face latérale du corps de la 12e vertèbre thoracique [5]. Ces dernières fibres peuvent être rejointes à leur insertion par les faisceaux des processus transverses lombaux – le plus souvent au niveau de L4 et L5, lorsque cela se produit.
La fonction exacte du carré des lombes est rendue difficile par sa structure irrégulière et inconstante. L’une des fonctions classiques de ce muscle serait de fixer la 12e côte lors de la respiration [2]. Cela correspond à l’insertion de la plupart de ses fibres (mais pas de toutes), sur la 12e côte. Cependant, la majorité des plus grosses fibres ancrent les processus transverses lombaux et la 12e vertèbre thoracique à l’ilium. Ces insertions indiquent que l’action principale de ce muscle devrait être la latéroflexion du rachis lombal. La force de ce muscle est cependant limitée par la taille de ses faisceaux et de leurs bras de leviers. En latéroflexion, le carré des lombes peut exercer un moment maximal d’environ 35 Nm [5].
Les faisceaux du carré des lombes sont des extenseurs potentiels du rachis lombal, car ils agissent derrière les axes de rotation sagittale des vertèbres lombales. Leur efficacité dans cette fonction est toutefois limitée à environ 20 Nm [5], ce qui équivaut à moins de 10 % du moment exercé par les muscles spinaux postérieurs.
Les limitations de ces forces perpétuent l’enigme sur la fonction réelle du carré des lombes.
MUSCLES SPINAUX LOMBAUX
Les muscles spinaux lombaux se trouvent en arrière du plan des processus transverses et exercent une action sur le rachis lombal. Ils comprennent les muscles s’attachant sur les vertèbres lombales et ayant une action directe sur le rachis lombal, ainsi que certains autres muscles qui, bien que non attachés aux vertèbres lombales, exercent néanmoins une action sur le rachis lombal.
• les muscles intersegmentaires courts : les interépineux et les faisceaux internes des intertransversaires ;
• les muscles polysegmentaires s’attachant aux vertèbres lombales : le multifidus et les composantes lombales du longissimus et de l’iliocostal ;
• les longs muscles polysegmentaires représentés par les composantes thoraciques du longissimus et de l’iliocostal qui, en général, ne s’insèrent pas aux vertèbres lombales, mais traversent la région lombale à partir des niveaux thoraciques pour s’insérer sur l’ilium et le sacrum.
Les descriptions des muscles spinaux proposées dans ce chapitre, en particulier celles du multifidus et des muscles érecteurs spinaux, diffèrent en grande partie de celles des manuels classiques. Ces muscles sont traditionnellement considérés comme ayant la même origine sur le sacrum et l’ilium et se dirigeant vers le haut pour s’insérer séparément sur les vertèbres dorsales, lombales et les côtes. Eu égard aux différentes études sur ces muscles [6–9], il est toutefois plus pertinent de les examiner en direction inverse, de haut en bas. Cela est non seulement plus cohérent avec le type d’innervation [9,10], mais explique aussi la particularité de certains muscles ainsi que l’aponévrose des muscles érecteurs spinaux, et révèle l’organisation biomécanique segmentaire de ces muscles.
Interépineux
Les interépineux lombaux sont des muscles pairs reposant de chaque côté du ligament interépineux et reliant les processus transverses des vertèbres lombales adjacentes (voir figure 9.2). Il en existe quatre paires dans la région lombale.
Intertransversaires médiaux
Les intertransversaires médiaux peuvent être considérés comme de vrais muscles spinaux, car contrairement aux intertransversaires latéraux, ils sont innervés par les rameaux dorsaux lombaux [3,10]. Ils ont pour origine le tubercule accessoire, le processus mamillaire attenant et le ligament mamillo-accessoire qui relie ces deux processus [11]. Ils s’insèrent sur la partie supérieure du processus mamillaire de la vertèbre du dessous (voir figure 9.2).
Une proposition alternative extrêmement attrayante serait que les intertransversaires (et peut-être aussi les interépineux) se comportent comme de nombreux capteurs proprioceptifs. Leur intérêt ne réside pas dans la force qu’ils peuvent exercer, mais dans les fuseaux musculaires qu’ils contiennent. Placés près de la colonne lombale, les intertransversaires pourraient contrôler les mouvements de la colonne et apporter un feedback influençant l’action des muscles environnants. Un tel rôle a été suggéré pour les intertransversaires cervicaux sur lesquels on a trouvé une grande densité de fuseaux musculaires [12–14]. Les muscles unisegmentaires de la colonne vertébrale ont en effet une densité de fuseaux musculaires deux à six fois supérieure aux muscles polysegmentaires plus longs. On pense de plus en plus que cela souligne la fonction proprioceptive de tous les petits muscles du corps humain [15,17].
Multifidus
Le multifidus est le plus important et le plus interne des muscles lombaux spinaux. Il est composé d’une série de faisceaux provenant des lames et des processus transverses des vertèbres lombales, et présente des insertions caudales en ordre régulier [9].
Les faisceaux les plus courts du multifidus sont les « fibres laminaires » provenant de l’extrémité caudale de la surface dorsale de chaque lame vertébrale ; ils s’insèrent deux niveaux plus bas sur le processus mamillaire de la vertèbre (figure 9.4A). Les fibres laminaires de L5 n’ont pas de processus mamillaire sur lequel elles peuvent s’insérer. Elles s’insèrent sur la région du sacrum qui se trouve juste au-dessus du premier foramen sacré postérieur. Les fibres laminaires peuvent être considérées équivalentes aux rotateurs thoraciques.
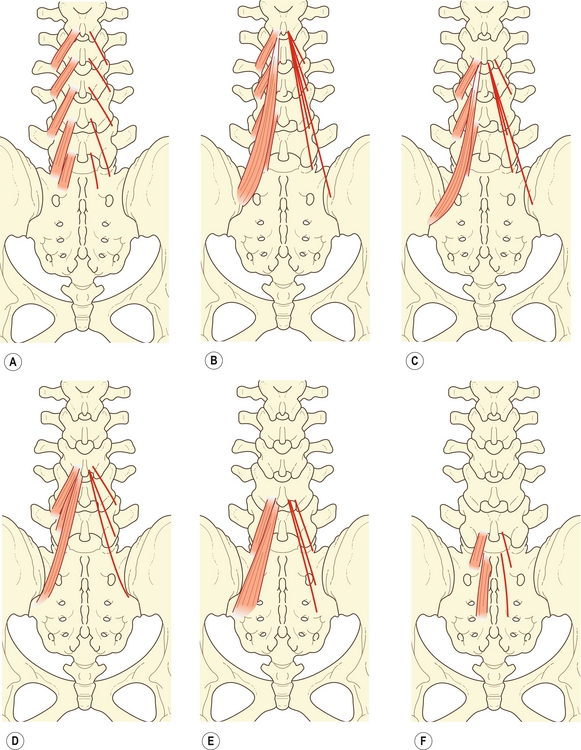
Figure 9.4 Composantes fasciculaires du multifidus. A : Fibres laminaires du multifidus. B–F : Faisceaux respectifs des processus épineux de L1 à L5.
Le faisceau de la base du processus épineux de L1 s’insère sur le processus mamillaire de L4, alors que celui du tendon commun s’insère sur les processus mamillaires de L5, S1 et sur l’épine iliaque postérosupérieure (figure 9.4B).
Le faisceau partant de la base du processus épineux de L2 s’insère sur le processus mamillaire de L5, alors que ceux du tendon commun s’insèrent sur le processus mamillaire de S1, sur l’épine iliaque postérosupérieure, et sur une région de la crête iliaque caudoventrale à l’épine iliaque postérosupérieure (figure 9.4C).
Le faisceau de la base du processus épineux de L3 s’insère sur le processus mamillaire du sacrum, alors que les faisceaux du tendon commun s’insèrent sur une région étroite s’étendant caudalement de l’expansion caudale de l’épine iliaque postérosupérieure au bord latéral du troisième segment sacré (figure 9.4D). Les faisceaux de L4 s’insèrent sur le sacrum et sur une région médiale à la région d’insertion de L3, mais latérale aux foramens sacrés postérieurs (figure 9.4E), alors que ceux de la vertèbre L5 s’insèrent sur une région médiale aux foramens sacrés postérieurs (figure 9.4F).
Il convient de noter que malgré les insertions de nombreux faisceaux du multifidus sur les processus mamillaires, quelques fibres profondes de ces faisceaux s’insèrent aux capsules des articulations zygapophysaires près des processus mamillaires [18] (voir chapitre 3). Ces insertions permettent au multifidus d’éviter que la capsule articulaire ne soit prise au piège à l’intérieur de l’articulation lors des mouvements exécutés par le multifidus.
La caractéristique clé de la morphologie du multifidus lombal provient de la disposition segmentaire de ses faisceaux. Chaque vertèbre lombale est dotée d’un groupe de faisceaux qui irradient du processus épineux et s’ancrent plus bas sur les processus mamillaires, la crête iliaque et le sacrum. Cette disposition laisse à penser que les fibres du multifidus sont disposées de telle manière que leur action principale est focalisée sur les processus épineux lombaux individuels [9]. Elles sont conçues pour agir de concert avec un seul processus épineux. Cette opinion est confirmée par le type d’innervation du muscle. Tous les faisceaux provenant des processus épineux d’une vertèbre donnée sont innervés par la branche médiane du rameau dorsal émergeant sous cette vertèbre [9,10] (voir chapitre 10). Par conséquent, les muscles qui agissent directement sur une vertèbre spécifique sont innervés par le nerf de ce segment.
Sur une vue postérieure, les faisceaux du multifidus ont une orientation oblique et caudolatérale. Leur ligne d’action peut donc être réduite à deux vecteurs : un grand vecteur vertical et un vecteur horizontal considérablement plus petit [7] (figure 9.5A).
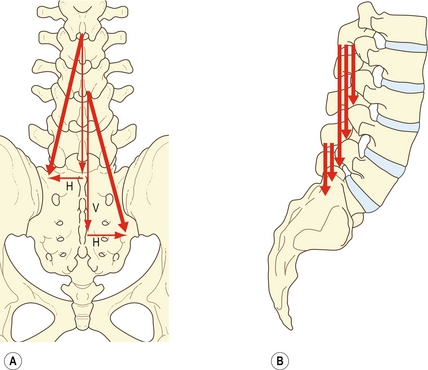
Figure 9.5 Forces vectorielles du multifidus. A : Sur une vue postéro-antérieure, la ligne de force oblique du multifidus peut être réduite en un vecteur vertical principal (V) et en un vecteur horizontal plus petit (H). B : Sur une vue latérale, les vecteurs verticaux du multifidus sont alignés perpendiculairement aux processus épineux.
Le petit vecteur horizontal laisse à penser que le multifidus pourrait tirer le processus épineux sur le côté et ainsi produire une rotation horizontale. Cependant, la rotation horizontale des vertèbres lombales est entravée par l’impaction des articulations zygapophysaires controlatérales. La rotation horizontale se produit après l’impaction des articulations, à condition qu’une force de cisaillement suffisante soit appliquée sur les disques intervertébraux (voir chapitre 7). La petite taille du vecteur horizontal du multifidus rend peu probable l’exercice d’une telle force de cisaillement sur le disque par l’action sur le processus épineux. Les études électromyographiques révèlent en effet que le multifidus est inconstamment actif en dérotation et qu’il est paradoxalement actif en rotation ipsilatérale et controlatérale [19]. On ne peut donc pas en déduire que la rotation est l’action principale du multifidus. Dans ce contexte, le multifidus est supposé se comporter uniquement comme un « stabilisateur » en rotation [18,19], mais les mouvements anormaux qu’il est censé stabiliser n’ont pas été déterminés (voir ci-après).
L’action principale du multifidus est mise en évidence par son vecteur vertical ; la projection latérale nous en apporte une meilleure compréhension (figure 9.5B). L’action de chaque faisceau du multifidus est pratiquement perpendiculaire à son processus épineux d’origine, sur tous les niveaux [7]. En utilisant le processus épineux comme levier, chaque faisceau est donc idéalement disposé pour produire une rotation sagittale postérieure de sa vertèbre. L’orientation perpendiculaire empêche cependant toute translation horizontale postérieure. Le multifidus ne peut donc qu’exercer la composante de « balancement » de l’extension du rachis lombal, ou ne contrôler que cette composante lors de la flexion.
Après avoir établi que le multifidus est principalement un rotateur sagittal postérieur du rachis lombal, il est possible d’expliquer son activité paradoxale lors de la rotation horizontale du tronc [7]. En premier lieu, il faut savoir que la rotation du rachis lombal est un mouvement secondaire. La rotation active du rachis lombal ne se produit que si le thorax est d’abord en rotation. Elle est donc secondaire à la rotation thoracique. Deuxièmement, il faut savoir qu’un muscle à deux vecteurs de force ne peut utiliser ces vecteurs séparément. Les deux vecteurs entrent en jeu lorsque le muscle se contracte. Le multifidus ne peut donc pas exercer une rotation axiale sans exercer simultanément une rotation sagittale postérieure bien plus importante.
Les principaux muscles produisant la rotation du thorax sont les muscles obliques abdominaux. La composante horizontale de leur orientation permet de faire pivoter la cage thoracique dans le plan horizontal et contribue ainsi à la rotation axiale au rachis lombal. Cependant, les muscles obliques abdominaux ont aussi une composante verticale. Leur contraction créera donc une rotation entraînant simultanément une flexion du tronc et du rachis lombal. Pour contrebalancer cette flexion et maintenir une pure rotation axiale, les extenseurs du rachis lombal doivent être recrutés, et c’est de cette façon que le multifidus se trouve associé à la rotation.
Le rôle du multifidus dans la rotation n’est pas de créer une rotation, mais de s’opposer à la force de flexion des muscles abdominaux en rotation. Le mouvement « anormal » stabilisé par le multifidus lors de la rotation est donc la flexion indésirable inévitablement produite par les muscles abdominaux [7].
Muscles érecteurs spinaux lombaux
Les muscles érecteurs spinaux lombaux sont situés latéralement au multifidus et forment le contour dorsolatéral proéminent des muscles spinaux de la région lombale. Ils sont composés de deux muscles, le longissimus thoracique et l’iliocostal lombal. En outre, chacun de ces muscles a deux composantes : une partie lombale composée de faisceaux provenant des vertèbres lombales, et une partie thoracique composée de faisceaux provenant des vertèbres thoraciques ou des côtes [6,8]. Ces quatre parties portent respectivement le nom de portion lombale du longissimus thoracique, portion lombale de l’iliocostal lombal, portion thoracique du longissimus thoracique, et portion thoracique de l’iliocostal lombal [8].
Dans la région lombale, le longissimus et l’iliocostal sont séparés l’un de l’autre par l’aponévrose lombale intermusculaire qui est le prolongement antéropostérieur de l’aponévrose des érecteurs spinaux lombaux [6,8]. Elle a l’aspect d’un feuillet plat de fibres de collagène se prolongeant rostralement sur 6 à 8 cm à partir de la partie interne de l’épine iliaque postérosupérieure. Elle est composée principalement des tendons caudaux des quatre faisceaux rostraux de la composante lombale du longissimus (figure 9.6).
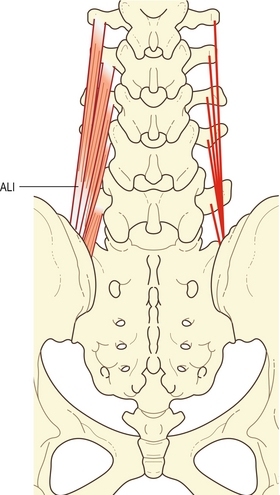
Figure 9.6 Fibres lombales du longissimus (longissimus thoracis pars lumborum). À gauche sont dessinés les cinq faisceaux du muscle complet. Représentation de la formation aponévrotique lombale intermusculaire (ALI) par les faisceaux lombaux. Sur la droite, les lignes indiquent les insertions et l’étendue des faisceaux.
Portion lombale du longissimus thoracique (longissimus thoracis pars lumborum)
La portion lombale du longissimus thoracique est composée de cinq faisceaux provenant chacun du tubercule accessoire et de l’extrémité médiale adjacente de la surface dorsale du processus transverse de la vertèbre lombale (voir figure 9.6).
Tous les faisceaux de L1 à L4 forment des tendons sur leurs extrémités caudales qui convergent pour former l’aponévrose lombale intermusculaire. Celle-ci s’insère pour finir sur une zone étroite de l’ilium, directement latérale à l’insertion du faisceau de L5. L’aponévrose lombale intermusculaire représente ainsi un tendon commun d’insertion, ou l’aponévrose du corps des fibres lombales du longissimus.
Chaque faisceau de la portion lombale du longissimus possède une orientation dorsoventrale et rostrocaudale [8]. La force de chaque faisceau peut donc être déterminée par un vecteur vertical et un vecteur horizontal dont la taille relative varie de L1 à L5 (figure 9.7A). Les forces relatives du longissimus diffèrent donc à chaque niveau segmentaire. En outre, la force du longissimus variera, dans son ensemble, selon la contraction unilatérale ou bilatérale du muscle.
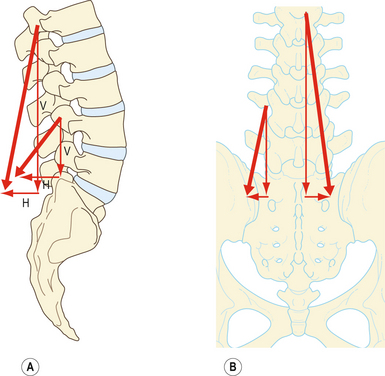
Figure 9.7 Vecteurs de force de la portion lombale du longissimus (longissimus thoracis pars lumborum). A : Sur une vue latérale, la ligne de force oblique de chaque faisceau du longissimus peut être représentée par un vecteur vertical (V) et un vecteur horizontal (H). Les vecteurs horizontaux des faisceaux des lombales basses sont plus grands. B : Sur une vue postéro-antérieure, la ligne de force des faisceaux peut être réduite en un vecteur vertical principal et en un vecteur horizontal beaucoup plus petit.
Le grand vecteur vertical de chaque faisceau repose latéralement à l’axe de flexion latérale et en arrière de l’axe de rotation sagittale de chaque vertèbre. Par conséquent, la contraction unilatérale du longissimus peut fléchir latéralement la colonne vertébrale, mais en fonctionnant bilatéralement, les divers faisceaux peuvent avoir la même action que le multifidus et provoquer une rotation sagittale postérieure de leur vertèbre d’origine. Toutefois, leurs insertions sur les tubercules accessoires et transverses reposent près des axes de rotation sagittale, et leur capacité à produire une rotation sagittale postérieure est moins efficace que le multifidus qui agit par l’intermédiaire des longs leviers des processus épineux [8].
Les vecteurs horizontaux du longissimus sont orientés vers l’arrière. Le longissimus est donc capable, en se contractant bilatéralement, de tirer les vertèbres lombales vers l’arrière. Cette force de translation postérieure peut rétablir la translation antérieure des vertèbres lombales se produisant lors de la flexion de la colonne lombale (voir chapitre 7). La capacité de translation postérieure est plus importante au niveau des lombales inférieures, où les faisceaux du longissimus acquièrent une orientation dorsoventrale plus importante (figure 9.7B).
Portion lombale de l’iliocostal lombal (iliocostalis lumborum pars lumborum)
La composante lombale de l’iliocostal lombal est constituée de quatre faisceaux superposés émergeant des vertèbres L1 à L4. Rostralement, chaque faisceau s’insère sur l’extrémité du processus transverse et, latéralement, sur les 2–3 cm de la couche intermédiaire du fascia thoracolombal (figure 9.8).
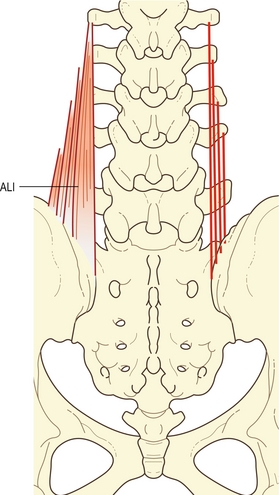
Figure 9.8 Fibres lombales de l’iliocostal (iliocostalis lumborum pars lumborum). Sur la gauche sont exposés les quatre faisceaux lombaux de l’iliocostal. Sur la droite, leurs surfaces et leurs insertions sont indiquées par des traits. ALI : formation aponévrotique lombale intermusculaire.
Le faisceau de L4 est le plus profond. Inférieurement, il s’insère directement sur la crête iliaque, juste latéralement à l’épine iliaque postérosupérieure. Ce faisceau est recouvert par le faisceau de L3 qui possède une attache semblable sur la crête iliaque, mais localisée plus dorsolatéralement. Dans l’ordre, L2 recouvre L3 et L1 recouvre L2, les insertions sur la crête iliaque devenant de plus en plus dorsales et latérales. Les faisceaux les plus latéraux s’insèrent sur la crête iliaque, médialement à l’attache du « raphé latéral » du fascia thoracolombal (voir ci-après). Une petite partie des fibres les plus médiales du muscle iliocostal participent à l’aponévrose intermusculaire lombale.
Malgré l’absence de description du faisceau L5 du muscle iliocostal dans la littérature, celui-ci est représenté par le « ligament » iliolombal. Chez les nouveau-nés et les enfants, la structure de ce « ligament » est considérée comme totalement musculaire [20] (voir chapitre 4). Le collagène remplace entièrement les fibres musculaires vers l’âge de 30 ans, donnant naissance au ligament iliolombal que l’on connaît [20]. En se fondant sur son site d’attache et son orientation correspondante, la bande postérieure du ligament iliolombal semblerait provenir du faisceau L5 de l’iliocostal, alors que la bande antérieure du ligament est dérivée du carré des lombes.
La disposition des faisceaux lombaux du muscle iliocostal est analogue à celle du longissimus, excepté leur position plus latérale. Comme pour le longissimus, leurs forces peuvent être réduites en vecteurs horizontaux et verticaux (figure 9.9A).
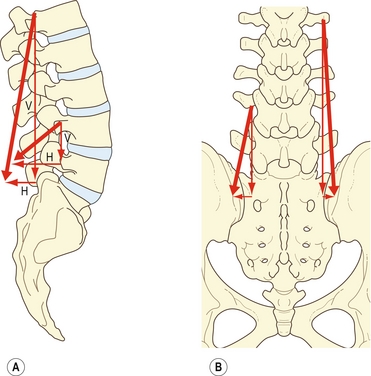
Figure 9.9 Forces vectorielles de la portion lombale de l’iliocostal lombal (iliocostalis lumborum pars lumborum). A : Sur une vue latérale, la ligne de force des faisceaux peut être réduite en vecteurs verticaux (V) et horizontaux (H). Les vecteurs horizontaux sont plus longs au niveau des lombales basses. B : Sur une vue postéro-antérieure, la ligne de force se réduit en un vecteur vertical et en un vecteur horizontal très petit.
Le vecteur vertical prédomine encore et la contraction bilatérale des faisceaux lombaux de l’iliocostal peut donc agir comme un rotateur sagittal postérieur (figure 9.9B). À cause de son vecteur horizontal, la translation postérieure sera exercée simultanément, particulièrement aux niveaux lombaux inférieurs où les faisceaux de l’iliocostal ont une orientation vers l’avant plus importante. En se contractant unilatéralement, les faisceaux lombaux de l’iliocostal peuvent se comporter comme des fléchisseurs latéraux des vertèbres lombales pour lesquels les processus transverses apportent des leviers considérables.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


