Chapitre 7
Bases biomécaniques
La biomécanique est une matière souvent décourageante et écrasante pour les étudiants en anatomie, à cause de son jargon et de son caractère mathématique. Certains concepts biomécaniques sont toutefois indispensables pour décrire et interpréter les mouvements ainsi que le vieillissement du rachis lombal. Il est par conséquent approprié d’examiner et de résumer ces concepts qui servent d’introduction aux chapitres abordant ces sujets.
MOUVEMENTS
Un os peut être sujet à deux types de mouvement : la translation et la rotation. Le principe de la translation réside dans le déplacement de tous les points d’un os dans une même direction et une même proportion (figure 7.1). La translation se produit à chaque fois qu’une force unique ou une force nette unique agit sur un os, et toute force qui tend à créer une translation est appelée force de cisaillement.
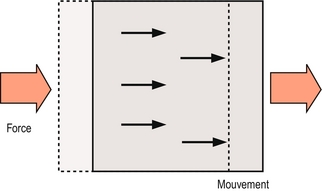
Figure 7.1 Translation. Une seule force nette entraîne le déplacement en parallèle de tous les points d’un corps dans la même direction et dans la même proportion.
La rotation est caractérisée par le déplacement parallèle de tous les points d’un os selon une trajectoire incurvée centrée sur un point fixe. Les points se déplacent dans une direction identique, mais dans différentes proportions selon leur distance radiale par rapport au point fixe appelé centre de rotation (figure 7.2). La rotation se produit lorsque deux forces non alignées agissent dans des directions opposées sur différentes parties d’un os, en créant ce que l’on appelle un couple de force (figure 7.2), et la force nette contribuant à la rotation est appelée couple. Selon les circonstances, le couple peut être le résultat de deux forces opposées qui peuvent toutes les deux être des forces musculaires, ou bien une force musculaire associée à une résistance ligamentaire, ou encore la force de gravité opposée soit à la réaction musculaire, soit à la résistance ligamentaire.
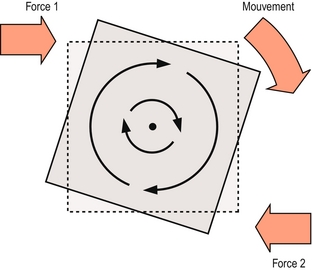
Figure 7.2 Rotation. Deux forces opposées non alignées (force de couple) entraînent le déplacement des points d’un corps autour d’un centre fixe.
Lorsque la rotation d’un os est examinée en trois dimensions, on peut voir que tous les points de l’os peuvent être assemblés en plans individuels reposant parallèlement à la direction du mouvement (figure 7.3). Dans chaque plan, les points se déplacent autour du centre localisé dans ce plan, et lorsque tous les centres de tous les plans sont alignés, ils dessinent une ligne droite formant ce qui est appelé l’axe de rotation osseux.
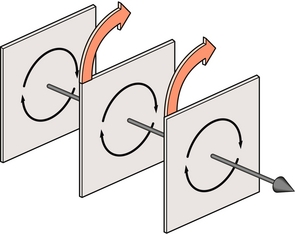
Figure 7.3 Lors de la rotation, les points de tous les plans d’un corps se déplacent autour d’un centre situé dans un de ces plans. La ligne formée par ces centres est l’axe de rotation du corps.
L’axe de rotation n’a pas de signification biologique particulière. Les points le long de l’axe de rotation n’ont aucune propriété biologique exceptionnelle. Un axe de rotation n’est qu’un phénomène mathématique créé par l’effet net des forces agissant sur un os. Pour toute rotation, on peut montrer qu’il existe une région où toutes les forces s’annulent et où aucune force nette n’est exercée ; il s’agit de l’axe de rotation. Cet axe reste immobile car aucune force nette n’est exercée sur lui. Au même moment, tous les points autour de l’axe sont soumis à une force nette et le mouvement se produit autour de cet axe stationnaire. Ainsi, la définition formelle de l’axe de rotation pourrait être celle d’une « zone immobile lors de l’action sur un os de deux ou plusieurs forces opposées non alignées ».
PLANS DES MOUVEMENTS
La translation et la rotation peuvent se produire dans l’une ou l’autre des directions opposées qui peuvent être définies de façon diverse selon les circonstances ou les usages. Par exemple, le mouvement peut être vers le haut ou vers le bas, en avant ou en arrière, dans le sens des aiguilles d’une montre ou dans le sens contraire, et selon certaines conventions positif (+) ou négatif (–). En outre, dans l’espace tridimensionnel, la translation et la rotation peuvent se produire dans n’importe lesquels des trois plans fondamentaux. En termes anatomiques, il s’agit du plan sagittal, frontal et horizontal (figure 7.4). Les rotations arrière ou avant sont des mouvements dans le plan sagittal, de même que les translations arrière ou avant. La flexion latérale est une rotation dans le plan frontal et la torsion est une rotation dans le plan horizontal. Un mouvement de glissement latéral dans le plan horizontal serait une translation horizontale, alors que des mouvements vers le haut ou vers le bas seraient décrits comme des translations frontales.
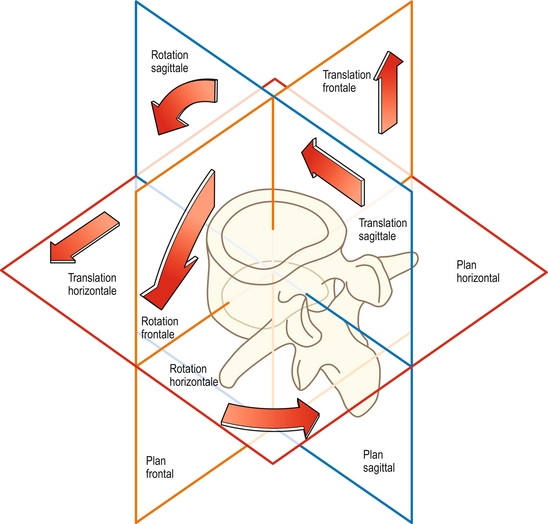
Les biomécaniciens préfèrent définir les mouvements par rapport aux trois axes imaginaires dessinés sur le corps vertébral [1,2]. L’axe X traverse obliquement le corps vertébral ; l’axe Y le traverse verticalement ; et l’axe Z le traverse d’arrière en avant (figure 7.5). Les mouvements peuvent ensuite être décrits selon leur position autour des axes ou le long de ces derniers. C’est ainsi que la translation sagittale est une translation le long de l’axe Z, les mouvements de glissement latéraux sont des translations le long de l’axe X, et les mouvements de haut en bas se font le long de l’axe Y. La flexion antérieure est une rotation autour de l’axe X, la flexion latérale est une rotation autour de l’axe Z, et les mouvements de torsion sont des rotations autour de l’axe Y. La clé de cette nomenclature repose sur les prépositions utilisées. Les translations sont des mouvements le long d’un des axes, et les rotations sont des mouvements autour d’un des axes.
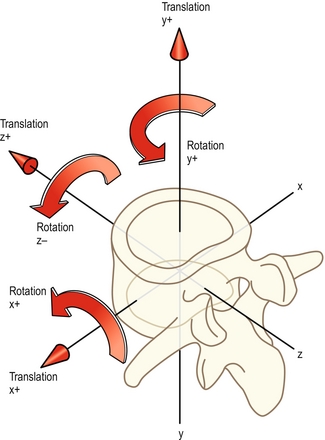
Les conventions des biomécaniciens ont pour avantage de définir précisément et sans ambiguïté les dimensions des mouvements. Cependant, les termes « X », « Y » et « Z » sont inhabituels et impersonnels pour tous, excepté pour ceux qui les utilisent régulièrement. Les adjectifs « sagittal », « frontal » et « horizontal » ont un peu plus de sens de par leur utilisation dans d’autres domaines de l’anatomie et sont les termes utilisés dans ce texte. Dans le système anatomique, les mouvements sont perçus comme se produisant dans le plan en question ou le long de celui-ci, sans savoir s’il s’agit d’une translation ou d’une rotation. Pour référence, l’équivalence des différents termes dérivés du système anatomique, ainsi que les usages des biomécaniciens et le vocabulaire familier sont présentés dans le tableau 7.1.
Tableau 7.1
Termes descriptifs du mouvement
| Système anatomique | Système biomécanique | Description familière |
| Translation sagittale antérieure | Translation Z + | Glissement vers l’avant |
| Translation sagittale postérieure | Translation Z – | Glissement vers l’arrière |
| Translation céphalofrontale | Translation Y + | Distraction longitudinale ou axiale |
| Translation caudofrontale | Translation Y – | Compression longitudinale ou axiale |
| Translation horizontale gauche | Translation X + | Glissement latéral gauche |
| Translation horizontale droite | Translation X – | Glissement latéral droit |
| Rotation sagittale antérieure | Rotation X + | Flexion en avant – « Flexion » |
| Rotation sagittale postérieure | Rotation X – | Flexion en arrière – « Extension » |
| Rotation frontale gauche | Rotation Z – | Flexion latérale gauche |
| Rotation frontale droite | Rotation Z + | Flexion latérale droite |
| Rotation horizontale gauche | Rotation Y + | Rotation axiale gauche |
| Rotation horizontale droite | Rotation Y – | Rotation axiale droite |
L’adjectif « axial » a été ajouté dans le tableau 7.1 en raison des difficultés à rendre compte des distinctions entre translations et rotations dans le plan horizontal et frontal, c’est-à-dire autour du long axe du corps. Ainsi, le terme « rotation axiale » remplace le terme « rotation horizontale » en référence à la rotation dans le plan horizontal, c’est-à-dire autour du long axe du corps. Le terme « translation axiale » remplace le terme « translation frontale » en référence aux mouvements vers le haut, vers le bas ou le long de l’axe du corps, et pour distinguer ce mouvement des translations obliques dans le plan horizontal qui sont décrites comme des translations horizontales ou latérales.
Les adjectifs « céphalique » et « caudale », qui signifient vers la tête et vers la queue, sont utilisés dans le tableau 7.1. Ces adjectifs sont peut-être lourds et inhabituels, mais ils sont précis et utilisables dans toutes les situations. Des expressions plus familières comme « vers le haut » ou « vers le bas » peuvent être employées pour les translations axiales en position debout, mais ne sont pas utilisables pour décrire rigoureusement les mouvements des vertèbres de patients qui pourraient être en décubitus. Pour surmonter cette difficulté, des termes plus familiers comme « distraction » ou « compression » sont habituellement utilisés à la place de translation axiale « céphalique » ou « caudale ». De même, le terme « flexion latérale » est plus commode et préféré au terme « rotation frontale ».
Dans ce texte, le terme rotation « sagittale » est strictement utilisé en référence aux mouvements rotatoires avant et arrière. Bien que les termes « flexion » et « extension » soient fréquemment utilisés pour décrire ce mouvement, ils ne sont pas suffisamment précis lorsqu’ils sont appliqués aux mouvements des vertèbres lombales individuelles. La flexion et l’extension ne sont pas de purs mouvements des vertèbres lombales, comme on le verra au chapitre 8 ; ces mouvements impliquent une combinaison de translation sagittale et de rotation sagittale. Les termes de flexion et d’extension peuvent être utilisés dans un sens général pour la flexion en avant et en arrière du rachis lombal, mais en ce qui concerne les mouvements des vertèbres individuelles, il devrait être entendu que les termes se réfèrent à la fois à la rotation sagittale et à la translation sagittale.
Ces définitions précises du mouvement ont un intérêt considérable. En premier lieu, le mouvement des vertèbres individuelles est souvent complexe, et aucun terme unique ne peut le décrire. Toutefois, il peut toujours être défini comme une combinaison de mouvements fondamentaux énumérée dans le tableau 7.1. De plus, chaque composante du mouvement du rachis lombal est exercée et entravée par différents mécanismes. Pour se rendre compte de la façon dont ces mécanismes entrent en jeu, chacun d’entre eux doit être analysé par rapport à la composante particulière du mouvement qu’il contrôle. Ce type d’analyse est entrepris au chapitre 8.
CONTRAINTE–DÉFORMATION
Il faut appliquer une force pour étirer une fibre de collagène. La fibre qui commence à être étirée résiste à l’allongement en générant une force de résistance en raison des liens chimiques entre les fibrilles de collagène, les molécules de tropocollagène, les fibres de collagène, et entre les fibres de collagène et les protéoglycanes (voir chapitre 2). Par convention, la force appliquée ou la force d’allongement porte le nom de contrainte appliquée, et le degré d’allongement de la fibre porte le nom de déformation. La contrainte est mesurée en unités de force (Newton), et la déformation se mesure en fraction ou en pourcentage d’augmentation de la longueur par rapport à la longueur d’origine. Ainsi, une fibre de longueur L0 étirée sur une longueur L1 subit une contrainte de L1/L0 ou L1/L0 × 100 %.
Des termes particuliers sont utilisés pour définir différents types de contraintes selon la direction dans laquelle la structure est déformée. Lorsqu’une structure est étirée longitudinalement, la force déformante porte le nom de tension, et la structure subit une déformation en tension. La contrainte de déformation est une compression si la structure est écrasée. Elle subit une déformation de compression. Cette dernière se mesure par la fraction ou le pourcentage de rétrécissement de la structure. Les forces qui entraînent le glissement d’une vertèbre sur l’autre portent le nom de forces de cisaillement, et la déformation se produisant sur le disque intervertébral intermédiaire est appelée déformation de cisaillement. La différence entre le cisaillement et la tension est que cette dernière s’applique conventionnellement aux forces le long du grand axe, alors que les forces de cisaillement sont appliquées sur cet axe. On dit qu’un objet se vrille lorsqu’il subit une torsion. Une force qui crée une torsion est un couple, et la déformation résultante est appelée déformation en torsion.
Au repos, les fibres de collagène isolées sont habituellement plissées et leur forme sinueuse porte le nom d’ondulation [3–6]. Lorsqu’une contrainte est appliquée à une fibre de collagène, le premier effet est l’aplanissement de cette ondulation. Cela nécessite peu d’énergie, car cette ondulation n’est pas maintenue par d’importants liens chimiques. Une fibre de collagène ondulée s’allongera donc en réponse à une force appliquée. Une fois l’ondulation supprimée, la fibre de collagène commence cependant à résister fortement à tout nouvel allongement. La contrainte tente de casser ces liaisons entre les fibrilles de collagène et les molécules de tropocollagène. De l’énergie est requise pour s’opposer à ces déformations et finalement rompre ces liaisons. L’allongement de la fibre de collagène requiert donc davantage de force. Les liaisons peuvent se rompre si une force suffisante est appliquée. Lorsque celle-ci se produit sur un grand nombre de liens, la fibre de collagène ne résiste plus à l’allongement et « rompt ». Une fois la fibre de collagène rompue, seules de petites forces sont requises pour la séparer de ses composantes fibrillaires et de ses molécules à présent non liées.
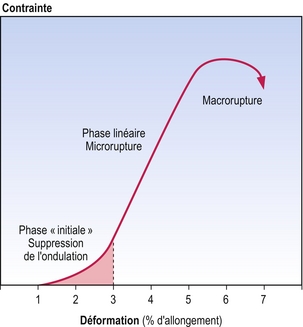
Le comportement mécanique des fibres de collagène sujettes à une contrainte peut être représenté graphiquement [7], comme sur la figure 7.6. Ces graphiques portent le nom de courbes de contrainte–déformation. La courbe présente trois régions principales. La première région, qui porte le nom de phase « initiale », reflète la phase de suppression de l’ondulation de la fibre de collagène. La seconde région linéaire se trouve au milieu de la courbe à forte inclinaison. Des calculs mathématiques ont révélé que la jonction entre la phase initiale et la région linéaire représente le point de suppression maximale de l’ondulation de la fibre, et le début de la contrainte d’étirement longitudinale de la fibre de collagène [6,7]. La région linéaire représente la phase de déformation et de cassure de quelques liens à l’intérieur des fibrilles de collagène et entre elles. Le sommet de la courbe représente la phase de rupture de la fibre de collagène, lorsqu’un nombre important de liens est rompu de manière irréversible. Une fois que la rupture s’est produite, la dernière partie de la courbe montre que l’allongement peut continuer avec des contraintes de moins en moins importantes.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


