Chapitre 2
Articulation intersomatique et disques intervertébraux
Une articulation pourrait être créée en plaçant simplement deux vertèbres successives l’une sur l’autre (figure 2.1A). Une telle articulation pourrait correctement supporter une charge et permettrait des mouvements de glissement entre les deux corps vertébraux. La platitude des surfaces articulaires ne permettrait cependant pas à l’articulation des mouvements de balanciers en flexion-extension ou en latéroflexion. Une ou deux modifications permettraient ces mouvements. La première consisterait à incurver les surfaces des corps vertébraux. La surface inférieure du corps vertébral pourrait par exemple être incurvée (comme les condyles fémoraux). La vertèbre supérieure d’une articulation intersomatique pourrait rouler en avant sur la surface plate du corps vertébral inférieur (figure 2.1B). Toutefois, cette modification compromettrait les capacités de port de charge et la stabilité de l’articulation intersomatique. La surface osseuse en contact avec la vertèbre inférieure serait réduite, et la propension de la vertèbre supérieure à rouler en avant et en arrière serait forte dès la mise en charge. Cette transformation serait par conséquent inadaptée si l’on veut préserver la capacité en charge et la stabilité de l’articulation intersomatique. Il convient de noter qu’il existe parmi quelques espèces animales, comme le poisson chez lequel la mise en charge n’est pas importante, une sorte d’énarthrose entre les corps vertébraux contribuant à la mobilité de la colonne vertébrale [1].
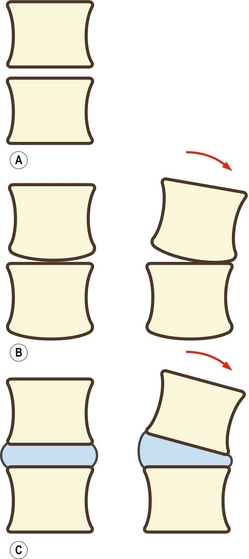
Figure 2.1 Agencements possibles d’une articulation intersomatique. A : Les corps vertébraux reposent directement l’un sur l’autre. B : L’ajout d’une courbure sur la partie inférieure d’une vertèbre permet de produire un mouvement de balancement. C : L’interposition de tissu mou écarte les corps vertébraux et permet de produire un mouvement de balancement.
La deuxième modification que l’on rencontre chez les humains et la plupart des mammifères consiste en l’interposition d’une couche solide mais déformable de tissu mou entre les corps vertébraux. Ce tissu mou se présente sous la forme d’un disque intervertébral. Ce disque a pour principal effet de séparer les deux corps vertébraux. L’espace entre les corps vertébraux permet à la vertèbre supérieure de s’incliner en avant sans que son bord distal rentre en contact avec le corps vertébral de la vertèbre inférieure (figure 2.1C).
Les prérequis biomécaniques qui en résultent pour le disque intervertébral sont de trois ordres. Il doit en premier lieu être assez solide pour supporter la charge, c’est-à-dire être capable de la transférer d’une vertèbre à une autre sans s’effondrer (ou s’écraser). Le disque doit deuxièmement être déformable pour s’adapter aux mouvements de balancier de la vertèbre sans compromettre outre mesure sa solidité. Troisièmement, il doit être assez solide pour ne pas être lésé lors des mouvements. La structure des disques intervertébraux doit donc être étudiée avec ces conditions préalables à l’esprit.
STRUCTURE DU DISQUE INTERVERTÉBRAL
Chaque disque intervertébral possède deux composantes de base : le nucléus pulposus central, entouré en périphérie par l’anulus fibrosus. Bien que le nucléus pulposus central soit tout à fait distinct de l’anulus fibrosus périphérique, il n’existe pas de frontière nette entre le nucléus et l’anulus à l’intérieur du disque. Les composantes périphériques du nucléus pulposus fusionnent plutôt avec les composantes les plus profondes de l’anulus fibrosus.
La troisième composante du disque intervertébral comprend deux couches de cartilage recouvrant la partie supérieure et inférieure du disque. Chacune porte le nom de plateau vertébral (figure 2.2). Les plateaux vertébraux séparent le disque des corps vertébraux adjacents et l’on peut se demander si ceux-ci sont des composantes discales strictes ou s’ils n’appartiennent pas plutôt à leur corps vertébral respectif. Dans cet ouvrage, il a été choisi de considérer le plateau vertébral comme une composante du disque intervertébral.
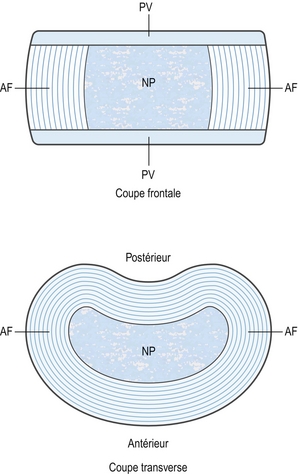
Figure 2.2 Structure basique d’un disque intervertébral. Le disque est constitué du nucléus pulposus (NP), entouré de l’anulus fibrosus (AF). Les deux sont pris en sandwich par les plateaux vertébraux cartilagineux (PV).
Nucléus pulposus
Le nucléus pulposus typique provenant de disques sains de jeunes adultes est une masse semi-liquide de matériel mucoïde ayant plus ou moins la consistance de pâte de dentifrice. Embryologiquement, le nucléus pulposus est un vestige de la notocorde (voir chapitre 12). Histologiquement, il est constitué de quelques cellules cartilagineuses et de quelques fibres irrégulières de collagène disséminées dans un milieu semi-liquide de substance fondamentale (voir ci-après). Biomécaniquement, la nature fluidique du nucléus pulposus lui permet de se déformer sous la pression, mais comme tout fluide, son volume ne peut être compressé. Lorsqu’il est soumis à des pressions multidirectionnelles, le nucléus aura tendance à se déformer et transmettra la pression appliquée dans toutes les directions. L’analogie avec un ballon rempli d’eau est pertinente. La compression du ballon le déforme. La pression dans le ballon augmente et étire ses parois dans toutes les directions.
Anulus fibrosus
L’anulus fibrosus est composé de fibres de collagène hautement organisées. Les fibres de collagène sont avant tout disposées en 10 à 20 feuillets [2,3] appelés lamelles (du latin lamella qui signifie petite feuille). Les lamelles sont disposées en anneaux concentriques entourant le nucléus pulposus (figures 2.2 et 2.3). Les lamelles sont plus épaisses vers le centre du disque [4]. Elles sont épaisses sur les versants antérieurs et latéraux de l’anulus, mais sont plus fines et plus serrées postérieurement. La partie postérieure de l’anulus fibrosus est par conséquent plus fine que le reste de l’anulus [2,5,6].
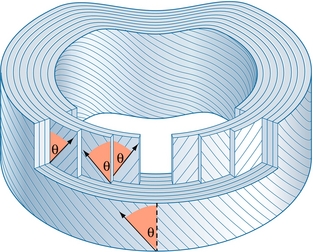
Figure 2.3 Structure détaillée de l’anulus fibrosus. Les fibres de collagène sont disposées en 10–20 lamelles concentriques circonférentielles. L’orientation des fibres à 65° alterne à chaque lamelle mais reste constante par rapport à la verticale (θ).
Les fibres de collagène sont disposées parallèlement à l’intérieur de chaque lamelle et passent de la vertèbre supérieure à la vertèbre inférieure. L’orientation des fibres de toutes les vertèbres est donc la même et leur angle est de 65–70° [7,8]. Toutefois, bien que l’angle soit le même, la direction de l’inclinaison alterne à chaque lamelle. Vues antérieurement, les fibres de chaque lamelle peuvent être orientées à 65° vers la droite, mais celles de la couche suivante seront orientées à 65° vers la gauche. Les fibres de la lamelle suivante seront à leur tour orientées à 65° vers la droite et ainsi de suite (figure 2.3). Une lamelle sur deux a exactement la même orientation. Ces chiffres ne constituent cependant qu’une orientation moyenne des fibres dans la portion intermédiaire des lamelles. Les fibres peuvent être plus ou moins fortement inclinées près de leur attachement par rapport au plan sagittal [4].
La description classique de l’anulus fibrosus présente les lamelles entourant complètement la circonférence du disque et formant un anneau complet. Cependant, il s’avère que ce n’est pas le cas. Chaque quadrant de l’anulus comporte près de 40 % de lamelles incomplètes, et le quadrant postérolatéral en comporte près de 50 % [4]. Une lamelle incomplète n’entoure pas entièrement la circonférence du disque. Les lamelles superficielles et profondes se rapprochent ou fusionnent près de leur extrémité terminale (figure 2.4). Les lamelles incomplètes semblent plus fréquentes dans la portion moyenne de l’anulus [9].
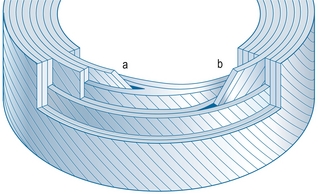
Plateaux vertébraux
Chaque plateau vertébral est composé d’une couche de cartilage d’environ 0,6 à 1 mm d’épaisseur [10–12] recouvrant la région du corps vertébral entourée par le listel marginal. Les deux plateaux vertébraux de chaque disque recouvrent entièrement le nucléus pulposus excepté en périphérie (figure 2.5). Histologiquement, le plateau est composé de cartilage hyalin et de fibrocartilage. Le cartilage hyalin se rencontre près du corps vertébral, en particulier chez les nouveau-nés et les disques jeunes (voir chapitre 12). Le fibrocartilage se rencontre près du nucléus pulposus et les plateaux vertébraux sont presque entièrement fibrocartilagineux chez les disques plus vieux (voir chapitre 13). Le fibrocartilage est formé par l’insertion de fibres collagènes de l’anulus fibrosus dans le plateau vertébral [6].
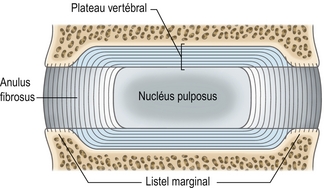
Figure 2.5 Structure détaillée d’un plateau vertébral. Les fibres de collagène des deux tiers internes de l’anulus fibrosus s’étendent à l’intérieur du plateau vertébral, formant ainsi sa composante fibrocartilagineuse. Les fibres périphériques de l’anulus sont ancrées dans l’os du listel marginal.
Les fibres de collagène des lamelles internes de l’anulus pénètrent le plateau vertébral et se retournent en son centre [3,13,14]. En suivant ces fibres sur leur longueur totale, on peut constater que le nucléus pulposus est entouré de tous côtés par une sphère de fibres de collagène ressemblant plus ou moins à une capsule. Cette « capsule » apparaît antérieurement, postérieurement et latéralement comme la lamelle la plus intime de l’anulus fibrosus, mais elle est absorbée supérieurement et inférieurement par les plateaux vertébraux (figure 2.5).
Les fibres collagènes des lamelles superficielles de l’anulus s’insèrent directement sur la partie osseuse du corps vertébral lorsque celui-ci est incomplet, comme sur le listel marginal [14] (figure 2.5). Dans leur forme originelle et dans des disques jeunes, ces fibres s’insèrent sur le plateau vertébral qui recouvre complètement les corps vertébraux du rachis lombal en croissance, mais ils sont secondairement absorbés par l’os lorsque le listel marginal s’ossifie (voir chapitre 12).
Les plateaux vertébraux sont solidement ancrés au disque intervertébral par l’intermédiaire de l’anulus fibrosus. Au contraire, les plateaux sont faiblement attachés aux corps vertébraux [13,14] et peuvent se détacher complètement de ceux-ci dans certains types de traumatismes rachidiens [15]. C’est pour cela et d’autres raisons morphologiques que les plateaux vertébraux sont considérés comme des constituants du disque intervertébral plutôt que comme des parties des corps vertébraux [10,12,13,16–18].
L’os sous-chondral est déficient sous une partie de la surface du plateau vertébral (à peu près 10 %) et des poches de cavité médullaire se juxtaposent à la surface du plateau ou pénètrent sur une courte distance à l’intérieur de celui-ci [11,19]. Ces poches facilitent la diffusion des nutriments à partir des vaisseaux sanguins vers l’espace médullaire et sont importantes pour la nutrition du plateau et du disque intervertébral (voir chapitre 11).
STRUCTURE DÉTAILLÉE DU DISQUE INTERVERTÉBRAL
Glycosaminoglycanes
Les glycosaminoglycanes (GAG) sont une classe chimique présente dans la plupart des tissus connectifs. On les trouve dans la peau, les os, le cartilage, les tendons, les valves cardiaques, les parois artérielles, le liquide synovial et l’humeur aqueuse de l’œil. Chimiquement, ce sont de longues chaînes de polysaccharides dont chacune est composée d’une séquence répétée de deux molécules appelées unités de répétition [20,21] (figure 2.6). Ces unités de répétition sont composées d’enchaînements de molécules de sucre et de molécules de sucre aminées. La nomenclature utilise le terme « glycosaminoglycane » (GAG) qui reflète bien les séquences « sucre–amine–sucre » dans leur structure.
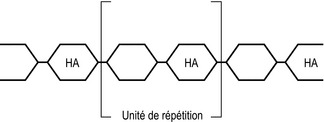
Figure 2.6 Structure moléculaire d’un mucopolysaccharide. La molécule est composée d’une chaîne de molécules de sucre, chacune étant composée d’un noyau de six carbones (hexose). Un sucre sur deux est une hexosamine (HA). La chaîne est une répétition de paires identiques d’hexose et d’hexosamine appelées unités de répétition.
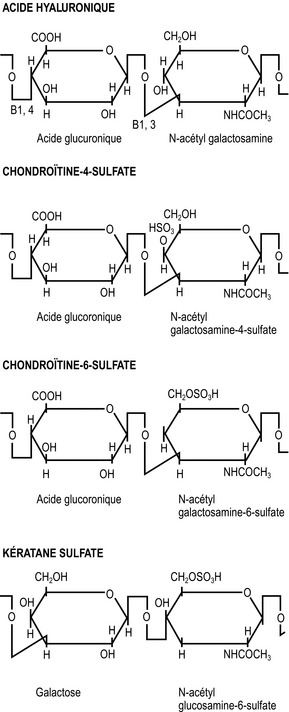
La longueur de chaque GAG varie mais est typiquement de 20 unités [21]. Chaque GAG est caractérisé par des molécules particulières qui composent sa séquence. Les GAG qui prédominent dans les disques intervertébraux humains sont la chondroïtine-6-sulfate, la chondroïtine-4 sulfate, le kératane sulfate et l’acide hyaluronique [22,23]. Les structures des unités de répétition de ces molécules sont présentées sur la figure 2.7.
Protéoglycanes
Les protéoglycanes sont de grosses molécules constituées de nombreux glycosaminoglycanes liés à des protéines. On les trouve sous deux formes élémentaires : les unités et les agrégats de protéoglycanes. Des unités de protéoglycanes sont créées lorsque plusieurs glycosaminoglycanes sont liés à une chaîne polypeptidique portant le nom de protéine centrale [22,24] (figure 2.8). Une simple protéine centrale peut transporter de six à 60 chaînes de polysaccharides [21]. Les glycosaminoglycanes sont liés à la protéine centrale par des ponts covalents nécessitant des molécules de sucre particulières [22,23]. Des agrégats de protéoglycanes se forment lorsque plusieurs unités de protéoglycanes se lient à une chaîne d’acide hyaluronique. Une simple chaîne hyaluronique peut lier de 20 à 100 unités de protéoglycanes [22]. Le lien entre les unités de protéoglycanes et l’acide hyaluronique est stabilisé par une relativement petite quantité de protéines appelées protéines de liaison (figure 2.8) [22].
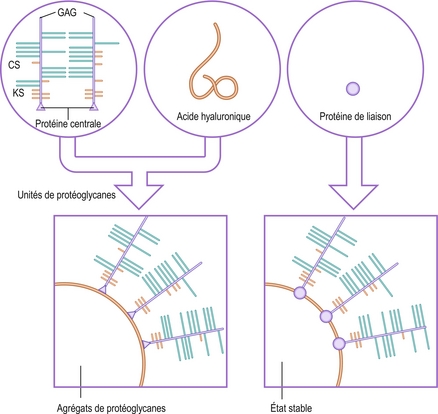
Figure 2.8 Structure des protéoglycanes. Les unités de protéoglycanes sont constituées de nombreux glycosaminoglycanes (GAG) liés à une protéine centrale. Les chaînes de kératane sulfate (KS) ont tendance à se trouver plus près de la tête de la protéine centrale. Des chaînes plus longues de chondroïtine sulfate (CS) sont attachées tout le long de la protéine centrale. Des agrégats de protéoglycanes se forment lorsque plusieurs unités de protéines se lient à une chaîne d’acide hyaluronique. Leur lien est stabilisé par une protéine de liaison.
Le protéoglycane essentiel du disque intervertébral ressemble à celui du cartilage articulaire et se nomme aggrécane [25]. Sa structure détaillée se trouve à la figure 2.9. Sa protéine centrale présente trois régions en hélices appelées domaines globulaires (G1, G2 et G3) et deux régions relativement rectilignes appelées domaines étendus (E1 et E2) [26]. Les glycosaminoglycanes sont surtout plus densément liés au domaine E1. Les chondroïtines sulfates se lient environ à trois quart de l’extrémité du domaine E2 (c’est-à-dire vers la terminaison carboxyle ou C-terminale de la protéine centrale) [22–24,26]. Le kératane sulfate se lie principalement au N-terminal du domaine E2 mais aussi sur les chaînes de chondroïtine sulfate [22–24,26,27]. Quelques chaînes de kératane sulfate se lient aussi au domaine E1.
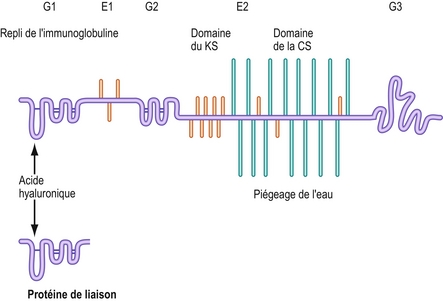
Figure 2.9 Structure de l’aggrécane. La protéine centrale présente trois domaines globulaires (G1, G2 et G3) et deux domaines étendus (E1 et E2). Le domaine E2 s’attache au kératane sulfate (KS) et à la chondroïtine sulfate (CS). Le domaine G1 est enroulé comme une immunoglobuline (Ig) de même que la protéine de liaison. Il est le site de liaison de la molécule d’aggrécane avec l’acide hyaluronique.
Le N-terminal de la protéine centrale porte le domaine E1 qui est replié comme une immunoglobuline. Une structure similaire est mise en évidence par la protéine de liaison. Ces structures hélicoïdales se lient à l’acide hyaluronique et permettent aux molécules d’aggrécanes de s’agglomérer [26]. Le domaine G1 n’acquiert sa structure qu’après le départ de la molécule d’aggrécane de la cellule qui l’a synthétisé [25]. Cela garantit une agglomération de la molécule d’aggrécane uniquement dans la matrice extracellulaire.
Les gros protéoglycanes qui s’agglomèrent à l’acide hyaluronique sont caractéristiques du cartilage hyalin et se trouvent parmi les disques intervertébraux immatures [23]. Ils sont riches en chondroïtine sulfate et transportent une centaine de ces chaînes d’un poids moléculaire d’environ 20 000. Ils transportent de 30 à 60 chaînes de kératane sulfate d’un poids moléculaire de 4000 à 8000 chacune [23]. Les protéoglycanes moyens et de grosses tailles qui ne s’agglomèrent pas à l’acide hyaluronique sont des protéoglycanes importants que l’on trouve dans les nucléus pulposus adultes [23].
In vivo, les unités et les agrégats de protéoglycanes sont spiralés pour former des molécules tridimensionnelles complexes ressemblant à des enchevêtrements de coton (figure 2.10). Physicochimiquement, ces molécules ont la capacité d’attirer et de retenir l’eau (mêmes propriétés absorbantes qu’une balle de coton). Une molécule de protéoglycane englobe un volume hydrophile appelé domaine [21].
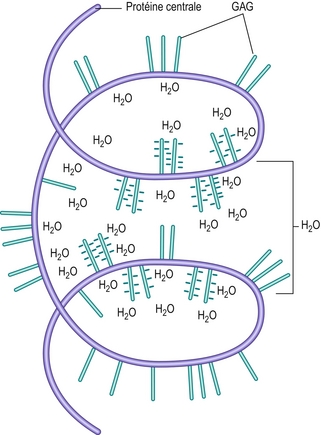
Figure 2.10 Aperçu d’une unité de protéoglycane hélicoïdale illustrant l’attraction de l’eau par les radicaux ioniques des glycosaminoglycanes (GAG) dans son « domaine ».
La capacité hydrophile d’une molécule de protéoglycane dépend de sa taille et de sa forme, mais la force hydrophile principale provient des radicaux carboxyles (COOH) et sulfates (SO4) des chaînes de glucosaminoglycanes (figure 2.7). Ces radicaux attirent l’eau électriquement et la capacité hydrophile d’un protéoglycane est proportionnelle à la densité structurelle de ces radicaux ioniques. À cet égard, les glycosaminoglycanes sulfatés sont plus hydrophiles que les autres mucopolysaccharides non sulfatés de taille similaire. En outre, il est évident que c’est parce que les chondroïtines sulfates possèdent des radicaux sulfatés et carboxyles sur leurs unités de répétition (figure 2.7) qu’ils ont deux fois plus de capacité hydrophile que le kératane sulfate qui, malgré son radical sulfaté, n’a pas de radical carboxyle. La capacité hydrophile de tout protéoglycane sera en grande partie dépendante de la concentration en chondroïtine sulfate dans sa structure [24].
Collagène
Le collagène est essentiellement composé de molécules de protéines filamenteuses. L’unité fondamentale du collagène est la molécule de tropocollagène, qui est elle-même composée de trois chaînes polypeptidiques enroulées hélicoïdalement et liées par des ponts hydrogènes (figure 2.11).
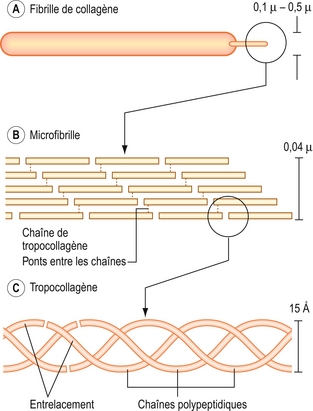
Figure 2.11 Structure du collagène. Une fibrille de collagène (A) est constituée de plusieurs microfibrilles de collagène (B). Chaque microfibrille est constituée de plusieurs chaînes de tropocollagène (C) maintenues ensemble côte à côte par des ponts covalents nécessitant des molécules d’hydroxylysine. Le tropocollagène est constitué de trois chaînes polypeptidiques enroulées l’une sur l’autre à la manière d’une hélice. Les chaînes de tropocollagène sont formées de chaînes polypeptidiques successives entrelacées et maintenues ensemble par des ponts électrostatiques entre leurs extrémités.
Le collagène est composé de molécules de tropocollagène arrangées côte à côte et à la suite l’une de l’autre. Lorsque quelques chaînes seulement sont accolées, la structure créée est une petite fibrille de collagène. Lorsque l’addition de plusieurs couches de chaînes de tropocollagène rend la structure plus épaisse, elle devient une grosse fibrille. L’agglomération de plusieurs grosses fibrilles forme une fibre de collagène. Les chaînes de tropocollagène à l’intérieur d’une fibre de collagène sont maintenues ensemble côte à côte par des ponts covalents associant une molécule d’hydroxylysine [28–30] (figure 2.11).
Il existe 11 types de collagène dans le tissu connectif [31]. Chaque type est génétiquement déterminé et diffère selon la nature chimique des chaînes polypeptidiques qui forment les molécules de tropocollagène trouvées dans la fibre de collagène et dans la microstructure de la fibre. Les différents types de collagène sont désignés par les chiffres romains de I à XI. Les types I, II, III, V et XI mettent en évidence la triple structure hélicoïdale typique décrite plus haut. Les types IV et VII sont de longues chaînes de molécules qui possèdent d’un côté une extension globulaire et dont la triple hélice est interrompue périodiquement par des segments non hélicoïdaux. Les types VI, VIII, IX et X sont des molécules beaucoup plus petites qui possèdent des segments hélicoïdaux discontinus ou identiques, porteurs d’extensions globulaires sur une ou deux de ces extrémités [31].
Les molécules de types I, II et III constituent la plus grande partie des fibres de collagène du corps humain. Les types I et II sont caractéristiques des tissus musculosquelettiques. Le tableau 2.1 indique leur distribution. Le collagène de type I est intrinsèquement de nature élastique et se trouve essentiellement dans les tissus soumis à des tensions et à des compressions. Le collagène de type II est de nature plus élastique et se trouve généralement dans les tissus exposés à la pression.
Tableau 2.1
Les types génétiques de collagène et leur distribution dans les tissus connectifs
| Type | Distribution |
| I | Peau, os, tendon, ménisques, dentine, anulus fibrosus |
| II | Cartilage, humeur vitrée, nucléus pulposus |
| III | Derme, cœur, vaisseaux sanguins, synoviale |
| IV | Membrane basale |
| V | Codistribué avec le type I |
| VI | Vaisseaux sanguins, viscères, muscle |
| VII | Membranes basales ectodermiques |
| VIII | Membrane de Descemet |
| IX | Cartilage, humeur vitrée |
| X | Plateaux épiphysaires |
| XI | Codistribué avec le type II |






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


