Chapitre 12
Embryologie et développement
Après 15 jours de développement, l’embryon humain a la forme d’un disque ovoïde et plat composé de deux couches de cellules : l’ectoderme dorsalement et l’endoderme ventralement (figure 12.1). L’ectoderme est la couche qui donnera principalement naissance à la peau et à la moelle spinale. L’endoderme formera le tube digestif [1].
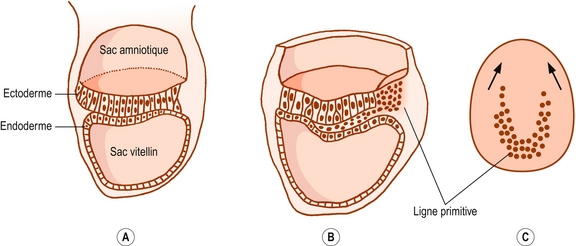
Figure 12.1 Développement du feuillet mésodermique chez de jeunes embryons. A : Coupe sagittale d’un jeune embryon composé uniquement d’ectoderme et d’endoderme. Le sac amniotique est disposé dorsalement à la plaque embryonnaire et le sac vitellin est suspendu au feuillet endodermique. B : Les cellules ectodermales à l’extrémité caudale d’un embryon de 15 jours se sont empilées pour former la ligne primitive qui donne naissance aux cellules mésodermales. C : Vue supérieure de l’embryon du schéma B montrant la migration vers l’avant des cellules mésodermiques des deux côtés du plan médian, sous le feuillet ectodermique.
Sur l’extrémité caudale de l’embryon, les cellules de l’ectoderme s’arrondissent et s’empilent en formant une saillie portant le nom de ligne primitive [1]. Les cellules de la ligne primitive migrent latéralement et vers l’avant. Elles s’infiltrent entre l’ectoderme et l’endoderme pour former une troisième couche embryonnaire appelée mésoderme (figures 12.1 et 12.2). Juste en avant de la ligne primitive se développe un épaississement appelé nœud de Hensen. Un cordon cellulaire appelé notocorde migre vers l’avant entre l’ectoderme et l’endoderme à partir de ce nœud (voir figure 12.2). Après environ 28 jours, la notocorde sépare complètement le plan médian de l’embryon [1] et induit la formation de la colonne vertébrale autour d’elle. Dorsalement à la notocorde, l’ectoderme forme le tube neural qui se différencie en cerveau et moelle spinale.
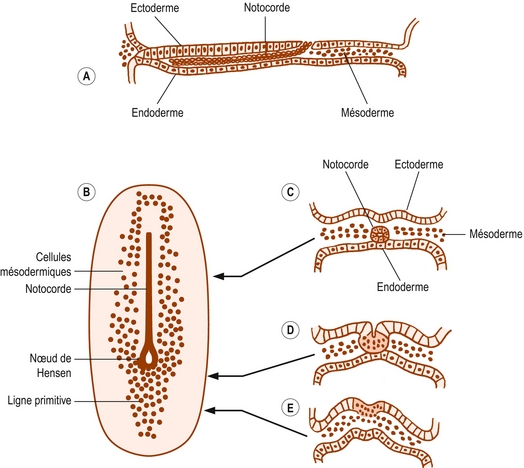
Figure 12.2 Développement mésodermique. A : Coupe sagittale d’un embryon montrant la notocorde qui s’est allongée vers l’avant entre l’ectoderme et l’endoderme et, derrière elle, le mésoderme de la ligne primitive. B : Vue d’en haut du même embryon montrant la notocorde et le mésoderme observés par l’intermédiaire de l’ectoderme au-dessus de l’embryon. C, D et E : Coupes transverses de l’embryon passant par la notocorde, le nœud de Hensen et la ligne primitive.
Le mésoderme de l’embryon s’épaissit de chaque côté de la notocorde pour former une masse longitudinale appelée mésoderme paraxial. Après le 21e jour de développement, le mésoderme paraxial commence à être marqué par des fissures transverses sur sa surface dorsale. Ces fissures séparent le mésoderme paraxial en segments appelés somites (figure 12.3). Les premiers somites apparaissent dans la région de la tête et d’autres apparaissent successivement caudalement. Un total de 42 à 44 somites est formé après environ 30 jours de développement embryonnaire [1].
Les fissures séparant les somites sont en réalité des indentations ; par conséquent, la segmentation qu’elles créent est apparente seulement le long de l’aspect dorsal du mésoderme paraxial. En profondeur, au-dessous de la surface de l’embryon, le mésoderme paraxial reste une masse longitudinale continue unique [2]. Le développement de chaque somite peut toutefois être suivi grâce à l’utilisation des fissures transverses comme repère.
Les 42 à 44 somites de l’embryon humain sont composés de 4 somites occipitaux, 8 cervicaux, 12 thoraciques, 5 lombaux, 5 sacrés et 8 à 10 coccygiens. Le premier somite occipital et les derniers sept à huit somites coccygiens régressent et ne donnent naissance à aucune structure durable [1]. Les trois somites restants sont associés à la formation de la région occipitale du crâne et de la langue. Les autres somites forment la colonne vertébrale et le tronc.
Les cellules somitiques sont originellement de nature épithéliale, mais se transforment progressivement en tissu lâche appelé mésenchyme (figure 12.4). En coupe transverse, le contour de chaque somite est à peu près triangulaire et présente des bords ventraux et dorsolatéraux ainsi qu’un bord neural faisant face au tube neural (figure 12.5).
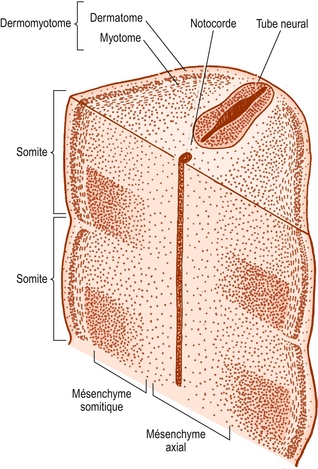
Figure 12.4 Coupes frontale et transverse combinées des somites d’un embryon. Le mésenchyme somitique s’est différencié en moitiés caudales denses et en moitiés craniales plus claires.
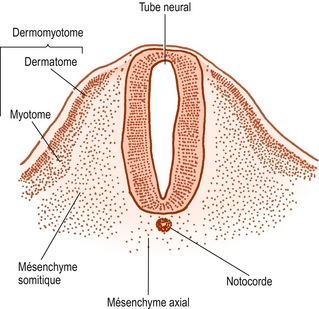
Figure 12.5 Coupe transverse d’un somite primitif montrant le lien entre le mésenchyme du tube neural et la notocorde ainsi que sa différenciation en mésenchyme somitique et en dermomyotome.
À l’intérieur du somite se développent deux groupes de cellules. Les cellules somitiques des régions ventrales et médiales se multiplient rapidement et forment une masse qui portait anciennement le nom de sclérotome, mais pour des raisons exposées ailleurs [2], le terme mésenchyme somitique est utilisé dans ce texte. Ces cellules sont exclusivement associées à la formation de la colonne vertébrale. Les cellules restantes le long du bord dorsolatéral du somite donnent naissance aux muscles et à la peau du tronc. Elles portent le nom de dermomyotome.
DESTINÉE DU MÉSENCHYME SOMITIQUE
Le mésenchyme somitique subit plusieurs changements qui entraînent finalement la formation d’un modèle rachidien primitif. Cette phase de développement de la colonne vertébrale est appelée phase mésenchymateuse.
La notocorde repose entre l’aorte ventralement et le tube neural dorsalement. Le tube neural est bordé par le mésenchyme somitique, mais au début celui-ci ne s’étend pas aussi loin que la notocorde médialement. La notocorde est entourée séparément par une colonne continue de mésenchyme très lâche appelé mésenchyme axial [2] (figure 12.5). La densité du mésenchyme axial augmente progressivement alors que ses cellules se multiplient et entourent la notocorde (figures 12.6 et 12.7). Pendant ce temps, une série d’événements indépendants se produisent dans le mésenchyme somitique.
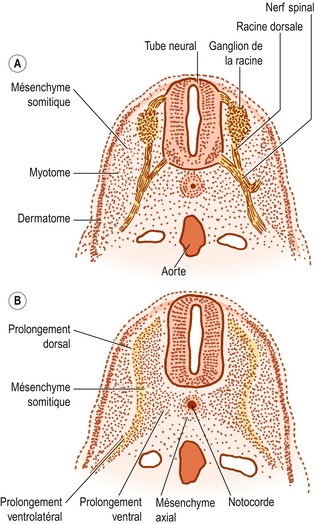
Figure 12.6 Coupes transverses de la moitié la moins dense d’un somite (A) et de la moitié caudale la plus dense (B). Sur la coupe A, le mésenchyme somitique entoure le nerf spinal en développement. Sur la coupe B, le mésenchyme axial entoure la notocorde. Le mésenchyme somitique a formé des prolongements dorsaux, ventrolatéraux et ventraux. (D’après Verbout [2].)
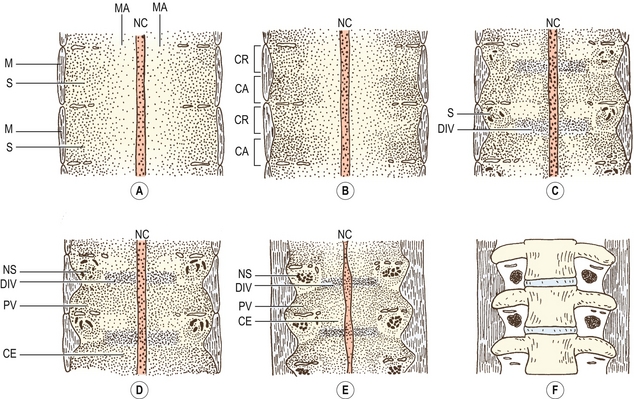
Figure 12.7 Aspect des coupes frontales de somites successifs montrant les étapes du développement des vertèbres lombales. A : Stade mésenchymateux primitif. M : myotome ; MA : mésenchyme axial ; NC : notocorde ; S : mésenchyme somitique. B : Les somites se sont différenciés en une moitié caudale dense (CA) et une moité crâniale moins dense (CR). C : Le mésenchyme s’est condensé autour du nerf spinal (NS) en développement. Le futur disque intervertébral (DIV) se caractérise par une zone de plus grande densité dans le mésenchyme axial, en face de l’extrémité inférieure de la moitié crâniale du somite. D : Le prolongement ventrolatéral (PV) du mésenchyme somitique s’étend entre deux myotomes successifs, et le prolongement ventral fusionne avec le mésenchyme axial pour former le centrum (CE). E : Les cellules mésenchymateuses se sont transformées en un modèle cartilagineux de la future vertèbre et la notocorde est extraite du corps vertébral. F : Localisation relative de la vertèbre osseuse définitive. (D’après Verbout [2].)
La densité des nucléus augmente dans la moitié caudale de chaque somite et leur donne une teinte plus foncée (voir figures 12.4 et 12.7B). La moitié crânienne du somite reste moins dense. Elle est envahie par le nerf spinal en développement (voir figures 12.6A et 12.7C). Le nerf croît latéralement pour envahir le dermomyotome, et les cellules de la moitié crânienne du somite s’organisent en couches concentriques autour du nerf au fur et à mesure que sa longueur et son épaisseur augmentent [2]. À terme, le développement du nerf occupe la plus grande partie de la moitié crânienne du somite qui donne lui-même naissance à un peu de tissu périneural. C’est la moitié caudale plus dense de chaque somite qui participe à la formation de la colonne vertébrale.
Dans la moitié caudale de chaque somite se développent deux prolongements : un prolongement dorsal et un prolongement ventrolatéral [2]. Le prolongement dorsal s’étend dorsalement pour entourer le tube neural et donne naissance à l’arc neural (voir figure 12.6B). Il porte donc aussi le nom de processus arqué. Le prolongement ventrolatéral s’étend latéralement et donne naissance à l’élément costal de la future vertèbre. Il est par conséquent appelé processus costal (voir figure 12.6B). Dans la région lombale, les éléments costaux de chaque vertèbre sont représentés sous la forme de processus transverses.
Au fur et à mesure que la densité du mésenchyme augmente, ses cellules prennent une orientation concentrique autour de la notocorde. Ces cellules vont former la plus grande partie du futur corps vertébral et la partie du corps qu’elles forment porte le nom de centrum (voir figure 12.7). En face de la moitié inférieure de la portion crâniale du somite adjacent, une zone de plus grande densité se développe dans le mésenchyme axial (voir figure 12.7). Cette zone constitue le précurseur du futur disque intervertébral [2].
Pendant le déroulement de ces phénomènes dans le mésenchyme axial, un troisième prolongement se développe dans le mésenchyme somitique. Ce prolongement, qui porte le nom de prolongement cordal ou ventral, s’étend vers la notocorde pour fusionner avec le mésenchyme axial juste caudal à la zone du futur disque intervertébral [2]. Le prolongement cordal relie de cette manière le mésenchyme somitique au centrum du corps vertébral. Le corps vertébral est finalement constitué du centrum et des parties terminales des prolongements cordaux de chaque côté.
Les prolongements dorsaux du mésenchyme somitique se poursuivent pour s’étendre autour des côtés du tube neural et juste latéralement à la racine du ganglion dorsal en développement. Les prolongements dorsaux des somites adjacents fusionnent les uns aux autres à l’endroit des futures articulations zygapophysaires [2]. À d’autres endroits, les arcs neuraux des segments adjacents sont liés par des condensations moins denses de mésenchyme qui donnent naissance aux ligaments de l’arc neural.
À ce stade du développement, la forme de la future vertèbre est représentée par le tissu mésenchymateux. Des condensations de mésenchyme axial ont entouré la notocorde et modelé le corps vertébral. Le futur disque intervertébral s’est condensé dans le mésenchyme axial à l’opposé de la moitié inférieure de la portion crâniale du mésenchyme somitique. La moitié crâniale de chaque somite s’est condensée autour du nerf spinal en développement et ne formera que du tissu périneural. La moitié caudale condensée du mésenchyme somitique a créé trois prolongements. Un prolongement ventral fusionne avec le mésenchyme axial sous le disque intervertébral, alors qu’un prolongement dorsal épouse le côté du tube neural. Les prolongements ventraux et dorsaux préfigurent le futur arc neural. Le prolongement ventrolatéral rayonne de chaque côté de l’arc neural en dessinant le futur processus transverse. À ce stade du développement, les prolongements dorsaux gauches et droits ne se joignent pas encore derrière le tube neural et ne sont unis que par une membrane [3,4]. L’arc neural est achevé dorsalement à un stade ultérieur du développement.
DESTINÉE DU DERMOMYOTOME
Originellement, deux types de cellules sont apparents dans le dermomyotome. Les cellules épithéliales recouvrent la surface dorsolatérale du somite et peuvent être identifiées en tant que dermatome. En profondeur de celles-ci reposent les cellules mésenchymateuses, connues collectivement sous le nom de myotome. Les cellules du dermatome perdent progressivement leur caractère épithélial et s’intègrent dans la masse myotomale, mais restent attachées au revêtement ectodermique, et donnent naissance au derme et aux tissus sous-cutanés [1]. Les cellules du myotome donnent naissance au tissu musculaire.
La masse myotomale reste localisée ventrolatéralement par rapport au mésenchyme somitique. En face de la moitié caudale condensée du somite, il est progressivement déplacé latéralement par le développement du prolongement ventrolatéral. En face de la moitié crâniale plus lâche du somite, il se distend dans sa direction, mais il est aussi dentelé par le nerf spinal en développement [2] (voir figure 12.7).
Alors que le nerf spinal se divise en un rameau ventral et dorsal au 40e jour environ de développement [4], le myotome se clive en deux parties [1]. La division se produit le long d’un plan représenté par les processus transverses en développement. Les deux portions sont séparées par un septum formant les futurs ligaments intertransversaires (figure 12.8). La portion dorsale du myotome, connue sous le nom d’épimère ou partie épaxiale, est innervée par le rameau dorsal du nerf spinal. La portion ventrale, connue sous le nom d’hypomère ou partie hypaxiale, est innervée par le rameau ventral du nerf spinal.
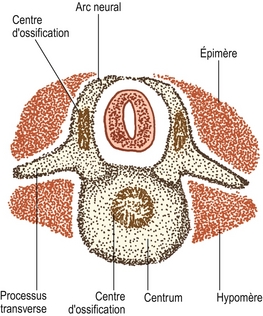
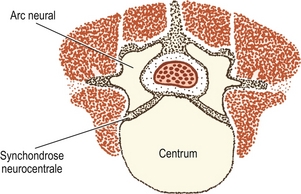
Figure 12.8 Coupe transverse d’une vertèbre lombale cartilagineuse montrant les centres d’ossification dans le centrum et les arcs neuraux ainsi que la disposition des myotomes dans les épimères et les hypomères.
Dans la région lombale, l’hypomère se développera en muscles ventraux aux ligaments intertransversaires. Les myotomes lombaux donnent naissance en grande partie aux muscles intertransversaires latéraux, au carré des lombes et au psoas. La plupart des muscles de la paroi abdominale se développent à partir des hypomères des somites thoraciques inférieurs, mais l’hypomère de L1 contribue aux parties inférieures de ces muscles.
Les épimères se divisent tout le long de la colonne vertébrale en une partie médiale et latérale [1,5], alimentées respectivement par les branches médiales et latérales des rameaux dorsaux. Dans la région lombale, la division médiale forme le muscle multifidus, alors que la division latérale forme les muscles iliocostal et longissimus.
CHONDRIFICATION
Alors que s’achèvent les modèles mésenchymateux des vertèbres, quelques cellules mésenchymateuses changent de nature et deviennent cartilagineuses, vers la 6e semaine de gestation [4], annonçant le début de la phase cartilagineuse du développement vertébral.
Deux centres de chondrification apparaissent au centre de chaque vertèbre. Ils fusionnent rapidement pour former un centre qui s’élargit en transformant en cartilage la totalité du centre [1]. Les centres de chondrification apparaissent aussi dans chaque moitié de l’arc neural. Ceux-ci s’étendent dorsalement sur le prolongement dorsal de chaque côté du mésenchyme somitique et se rejoignent à l’arrière du tube neural pour compléter l’arc neural. Un processus épineux cartilagineux se développe dorsalement à l’endroit de la jonction. Les centres de l’arc neural s’étendent aussi latéralement pour transformer en cartilage le processus transverse et pour fusionner avec le centre de chondrification ventralement, le long du prolongement ventral du mésenchyme somitique.
Le modèle cartilagineux de la future vertèbre est formé à la suite de ces phénomènes, mais ces modèles cartilagineux commencent à être remplacés par les vertèbres osseuses définitives lors même de la chondrification de la colonne vertébrale (voir figure 12.8).
OSSIFICATION
L’ossification est la troisième phase de développement de la colonne vertébrale. Elle commence lors de la 9e à la 10e semaine de vie intra-utérine [6], mais ne s’achève pas avant l’adolescence. Le premier processus d’ossification est appelé ossification primaire et se produit là où les vaisseaux sanguins envahissent les modèles cartilagineux des futures vertèbres.
Les arcs neuraux cartilagineux sont envahis par derrière pour former le centre d’ossification primaire dans chaque moitié de l’arc neural (voir figure 12.8). Le corps vertébral cartilagineux est envahi de vaisseaux sanguins par l’intermédiaire de ses surfaces antérieures et postérieures. Quelques experts maintiennent que ces deux séries de vaisseaux sanguins donnent naissance respectivement à des centres d’ossification dorsaux et ventraux séparés, qui fusionnent rapidement pour former un centre d’ossification unique au milieu du futur corps vertébral [7] ; mais d’autres affirment que ce phénomène n’est qu’une variante ne se produisant que dans 5 % des cas [8,9]. Une autre variante consiste en deux centres reposant latéralement l’un à l’autre, mais la variété la plus fréquente est celle qui contient un centre unique [8].
Le début de l’ossification diffère selon le niveau vertébral et la partie à ossifier. Les centres d’ossification primaires des arcs neuraux apparaissent d’abord au niveau cervicothoracique, suivis des cervicales hautes et du niveau thoracolombal. Les centres des arcs neuraux apparaissent ensuite progressivement dans les directions crâniale et caudale à partir de ces niveaux [10]. Les centres primaires des corps vertébraux apparaissent d’abord au niveau des thoraciques basses et des lombales hautes, puis apparaissent progressivement à des niveaux supérieurs et inférieurs [10]. Les centres d’ossification s’établissent de cette manière dans les corps et les arcs neuraux des vertèbres lombales, vers la 12e–14e semaine de gestation.
Dans le centrum du corps vertébral, le centre d’ossification primaire s’étend radialement et en direction des disques intervertébraux, au-dessus et en dessous. Il atteint l’aspect antérieur du centrum après environ 22 semaines de vie anténatale et l’aspect postérieur après environ 25 semaines [11] ; mais l’ossification n’atteint pas la surface supérieure et inférieure du corps vertébral, qui reste cartilagineux et forme les cartilages de croissance du corps vertébral. Dans les arcs neuraux, l’ossification s’étend dans toutes les directions à partir du centre primaire ; ventralement en direction du corps vertébral, latéralement vers le processus transverse, et dorsalement autour du tube neural.
À la naissance, les vertèbres lombales ne sont pas encore complètement ossifiées (figures 12.9 et 12.10). La plus grande partie du centrum est ossifiée. Sur des radiographies de profil, il a l’apparence d’un bloc osseux ovoïde avec des surfaces supérieures et inférieures convexes [12–14]. De nombreux canaux vasculaires pénètrent les côtés antérieurs et postérieurs du centrum [7], qui ont l’apparence de surfaces translucides sur les radiographies rachidiennes de nouveau-nés [14]. Les surfaces supérieure et inférieure du corps vertébral sont encore recouvertes de plaques épaisses de cartilage, et la hauteur combinée de ces plateaux et du disque intervertébral est approximativement la même que la hauteur des corps vertébraux lombaux ossifiés [12–14]. Les pédicules et les parties proximales des lames et des processus transverses sont ossifiés, mais les processus épineux et les parties distales des processus transverses sont encore cartilagineux. Les processus articulaires sont en grande partie ossifiés, mais leurs extrémités distales restent cartilagineuses.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


